Karoliina Hämäläinen  ,
Teemu Tahvanainen,
Kaisa Junninen
,
Teemu Tahvanainen,
Kaisa Junninen
Kuolleet puut ja niiden kääpälajisto lehdoissa
Hämäläinen K., Tahvanainen T., Junninen K. (2018). Kuolleet puut ja niiden kääpälajisto lehdoissa. Metsätieteen aikakauskirja vuosikerta 2018 artikkeli 10091. https://doi.org/10.14214/ma.10091
-
Hämäläinen,
Itä-Suomen yliopisto, Metsätieteiden osasto, Joensuu
Sähköposti
karoham@uef.fi
- Tahvanainen, Itä-Suomen yliopisto, Ympäristö- ja biotieteiden laitos, Joensuu Sähköposti teemu.tahvanainen@uef.fi
- Junninen, Metsähallitus, Eräpalvelut, Joensuu Sähköposti kaisa.junninen@metsa.fi
Vastaanotettu 3.12.2018 Hyväksytty 5.12.2018 Julkaistu 11.12.2018
Katselukerrat 32039
Saatavilla https://doi.org/10.14214/ma.10091 | Lataa PDF

Seloste artikkelista Hämäläinen K., Tahvanainen T., Junninen K. (2018). Characteristics of boreal and hemiboreal herb-rich forests as habitats for polypore fungi. Silva Fennica vol. 52 no. 5 article id 10001. https://doi.org/10.14214/sf.10001
Tutkimuksen tausta ja materiaalit
Boreaalisella vyöhykkeellä lehtoja pidetään metsien kasvupaikkatyypeistä lajistollisesti monimuotoisimpina ja siksi myös luonnonsuojelullisesti erityisen arvokkaina. Noin puolet Suomen uhanalaisista metsälajeista elääkin ensisijaisesti lehdoissa, vaikka lehtojen osuus maan pinta-alasta on alle prosentin.
Intensiivinen metsätalous on muuttanut monia boreaalisten metsien ominaispiirteitä viimeisten vuosikymmenten aikana, myös lehdoissa. Yksi merkittävimmistä muutoksista on raju lasku kuolleen puun määrässä. Kuolleesta puusta riippuvaisten lajien elinympäristöt ovat sekä määrällisesti vähentyneet että laadullisesti heikentyneet. Onkin esitetty, että Suomessa esiintyvistä 4000–5000:sta kuolleessa puuaineksessa elävästä lajista jopa yli puolet katoaisi talousmetsistä (Siitonen 2001).
Eräs keskeisimmistä kuolleessa puussa elävistä eliöryhmistä on käävät. Tutkimustietoa lehtojen käävistä on kuitenkin paljon vähemmän kuin kangasmetsien lajistosta. Tässä tutkimuksessa selvitettiin, miten substraatti- ja metsikkötason muuttujat vaikuttavat boreaalisten ja hemiboreaalisten lehtojen kääpälajistoon. Lisäksi tarkasteltiin, onko yleisten lajien ja Punaisen listan lajien (Kotiranta ym. 2010) vasteissa eroa.
Tutkimusaineistoon kuului 71 luonnontilaista tai luonnontilaisen kaltaista lehtokuviota (metsikköä) pohjoisboreaalisen vyöhykkeen eteläpuolelta. Kultakin metsäkuviolta tai metsäkuvion osalta kartoitettiin kaikki läpimitaltaan vähintään 10 cm lahopuukappaleet (kuolleen puun kappaleet) ja näiltä löydetyt käävät. Kartoitetun alueen pinta-ala laskettiin jälkikäteen GPS-jäljen perusteella. Lahopuukappaleista määritettiin puulaji, läpimitta, lahoaste viisiportaisella asteikolla ja puun tyyppi (maapuu, kanto, pökkelö, oksa, jne.).
Kääpälajistoa kuvaavina vastemuuttujina käytettiin kääpien lajimäärää, runsautta, Shannonin diversiteetti-indeksiä, Punaisen listan lajien lajimäärää ja asutettujen puiden osuutta kaikista lahopuista. Selittäviin muuttujiin kuuluivat kartoitetun alueen pinta-ala, pohjoiskoordinaatti, lahopuiden kokonaislukumäärä, lahopuiden lukumäärä hehtaaria kohti, suurikokoisten (läpimitta > 30 cm) lahopuiden lukumäärä hehtaaria kohti ja lahopuun diversiteetti.
Tulokset
Aineistoon kuului kaikkiaan 4797 lahopuukappaletta, joilta kertyi 2832 havaintoa 101 kääpälajista. Lehtipuilla kasvoi 70 ja havupuilla 50 lajia. Vain yhdeltä puulajilta havaittuja lajeja oli yhteensä 39. Eniten yhden puulajin kääpälajeja, 16 lajia, löytyi kuuselta. Punaisen listan lajeista kertyi 30 löytöä 12 lajista. Lahopuukappaleita oli keskimäärin 96,2 (SD ±77,2) hehtaaria kohden, mutta lukumäärä vaihteli huomattavasti. Keskimäärin 59 % (SD ±31 %) lahopuukappaleista oli lehtipuita, mutta yksittäisistä puulajeista kuusi oli kaikkein runsain. Lahopuukappaleista yli 60 % kuului pienimpään läpimittaluokkaan (10–19 cm). Tutkituiden metsäkuvioiden koko oli keskimäärin 0,91 hehtaaria (SD ±0,55 ha).
Metsikkötason muuttujista voimakkaimmin kääpien lajimäärän kanssa korreloivat lahopuun diversiteetti ja alueen pohjoisuus. Suurempi lahopuun diversiteetti kytkeytyi suurempaan lajimäärään, kun taas kasvavan pohjoiskoordinaatin ja kääpien lajimäärän suhde oli negatiivinen. Yhden selittäjän lineaarisen regressiomallin mukaan lahopuun diversiteetin vaihtelu selitti 45,5 % ja pohjoiskoordinaatti 16,6 % kääpien lajimäärän vaihtelusta. Lineaarinen monimuuttujaregressiomalli, jossa selittävinä muuttujina oli edellä mainittujen lisäksi myös suurten lahopuiden lukumäärä, selitti kokonaisuudessaan 52 % kääpien lajimäärän vaihtelusta. Punaisen listan kääpälajien lukumäärän vaihtelua selitti tilastollisesti merkitsevästi ainoastaan suurikokoisen lahopuun määrä, mutta mallin selitysaste oli varsin alhainen, 6,8 %.
Substraattitasolla korostui suurikokoisen lahopuun merkitys sekä kääpien lajimäärälle että runsaudelle. Lajien kertymäkäyrät olivat huomattavasti jyrkempiä läpimitaltaan vähintään 20 cm:n kuin läpimitaltaan 10–19 cm:n lahopuukappaleilla. Myös kääpien runsaus oli yleensä alhaisempi läpimitaltaan pienikokoisessa (10–19 cm) lahopuussa kuin suurikokoisessa (>30 cm), mutta ero oli tilastollisesti merkitsevä ainoastaan koivulahopuulla (Betula spp.).
Kaksiulotteisessa NMDS-ordinaatiossa, jossa tutkimusalueiden kääpäyhteisöjä tarkasteltiin puulajikohtaisesti, huomattavin ero yhteisöissä oli havu- ja lehtipuiden välillä. NMDS-ordinaation avulla tarkasteltiin myös kääpäyhteisöjä yleisesti ja ordinaatioon sovitettiin lajien NMDS-pisteiden kanssa tilastollisesti merkitsevästi (p < 0,05) korreloivat ympäristövektorit. Ympäristövektoreista tilastollisesti merkitseviä olivat pohjoisuus ja pinta-ala, sekä koivun, muiden lehtipuiden, jalojen lehtipuiden ja kuusen lahopuukappaleiden runsaus (Kuva 1).
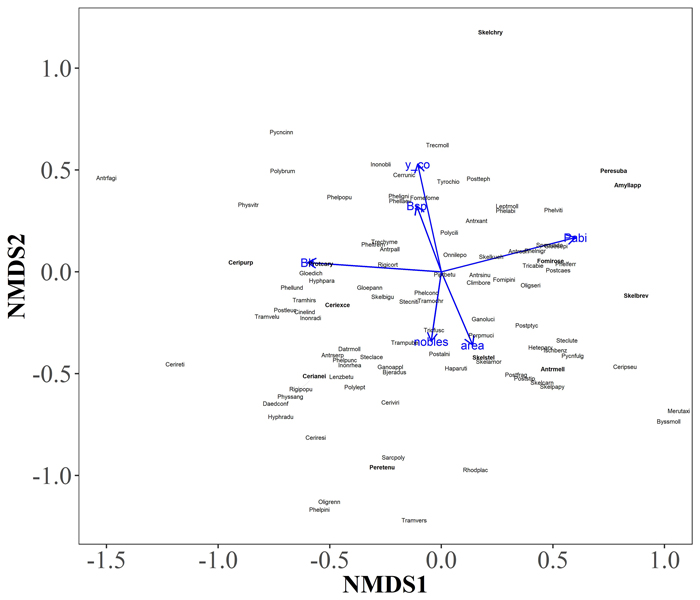
Kuva 1. Kaksiulotteinen NMDS-ordinaatio lehtojen kääpälajeista ja ordinaatioon sovitetut, lajipisteiden kanssa tilastollisesti merkitsevästi (p < 0.05) korreloivat ympäristövektorit. Pabi, kuusi Picea abies (r = 0.39); Blt, muut lehtipuut paitsi koivu ja haapa (r = 0.35); y_co, pohjoiskoordinaatti (r = 0.34); area, kartoitettu pinta-ala (r = 0.16); Bsp, koivu Betula spp. (r = 0.13); nobles, jalot lehtipuut (Tilia cordata, Acer platanoides ja Corylus avellana) (r = 0.13). Puulajivektorit kuvaavat kunkin lajin puukappaleiden määrää hehtaaria kohti. Uhanalaisten lajien nimien lyhenteet on vahvennettu. Katso isompi kuva uudessa ikkunassa.
Tulosten tarkastelu ja johtopäätökset
Tutkimusalueiden lahopuuprofiilin puulajisto kertoo varsin tyypillisestä boreaalisten lehtojen puustosta, jossa latvuskerros on kuusi-lehtipuu -sekametsää. Lehtipuiden osuus tutkimusalueiden lahopuuprofiilissa oli lähes kaksinkertainen verrattuna muissa tutkimuksissa mitattuihin vanhoihin kuusivaltaisiin kangasmetsiin (Siitonen ym. 2000).
Kuolleen puun suuri määrä ja diversiteetti lisäsivät lehtolaikun kääpien lajimäärää, runsautta ja diversiteettiä. Sekä korrelaatioanalyysin että lineaaristen regressiomallien tulokset viittaavat siihen, että lahopuun diversiteetti kuvaa tehokkaasti eri kääpälajeille tarjolla olevien ekolokeroiden määrää, ja että kuolleen puuston monimuotoisuus on ensisijaisen tärkeää monimuotoiselle kääpälajistolle.
Myös substraattitasolla tuloksissa korostui monipuolisen puulajiston ja suurikokoisen lahopuun merkitys. Ordinaatiossa kääpäyhteisöt erosivat erityisesti lehti- ja havupuiden välillä, ja suuri osa kääpälajeista tavattiin vain tietyn puulajin lahopuukappaleilta. Sama lukumäärä suurikokoista lahopuuta (>30 cm) elätti huomattavasti suuremman joukon kääpälajeja kuin sama määrä pienikokoista lahopuuta. Suurikokoisen lahopuun merkitys sekä kääpälajien lukumäärälle että yksilöiden runsaudelle on laajalti tiedetty (Junninen ja Komonen 2011). Tässä tutkimuksessa havaittiin, että lahopuun koon merkityksessä on eroja eri puulajien kesken, ja suurin ero kääpien runsaudessa löytyi läpimitaltaan pienten ja suurten koivulahopuiden väliltä.
Suurikokoisten kuolleiden puiden määrä osoittautui merkittävimmäksi muuttujaksi myös Punaisen listan kääville. Uhanalaiset ja silmälläpidettävät lajit ovat usein erikoistuneita käyttämään tietynlaisia substraatteja, kuten järeitä kuusi- ja mäntymaapuita. Tämän vuoksi punaisen listan lajeille on merkittävämpää se, onko metsäkuviolla riittävä määrä juuri niiden tarvitsemaa substraattia, eikä niinkään kuolleiden puiden kokonaismäärä tai diversiteetti. Tutkituista lehdoista löytyi verraten vähän Punaisen listan lajeja verrattuna aiempiin tutkimuksiin vanhoista kuusivaltaisista metsistä. Yksi mahdollinen syy tähän on lehtojen pienialaisuus ja pirstaloituneisuus, sillä Punaisen listan lajien on osoitettu reagoivan sopivan substraatin määrään paitsi metsikön sisällä myös maisematasolla (Nordén ym. 2013). Lisäksi monet lehdoissa elävät kääpälajit kasvavat suhteellisen yleisillä lahopuutyypeillä, kuten pienikokoisella lehtilahopuulla, eikä niitä siksi luokitella Punaisen listan lajeihin lajien uhanalaisuusarvioinneissa. Tämän seurauksena kääpälajistoltaan monipuolissakin lehtipuustoisissa lehdoissa on usein vähemmän Punaisen listan lajeja kuin kuusivaltaisissa vanhoissa metsissä.
Myös ilmastolliset tekijät näyttävät vaikuttavan lehtojen kääpälajistoon. Huolimatta siitä, että lahopuun diversiteetti ei korreloinut alueen pohjoisuuden kanssa, oli pohjoisuudella merkittävä negatiivinen vaikutus sekä kääpien lajimäärään, diversiteettiin että kääpien asuttamien lahopuukappaleiden osuuteen. Yksi mahdollinen syy tähän on lyhyt kasvukausi, joka saattaa vaikuttaa myös kääpien itiöimiskauden pituuteen.
Boreaalisella vyöhykkeellä lehtoja pidetään biodiversiteetin keskittyminä ensisijaisesti niiden rehevyyden ja täten rikkaan kasvilajiston vuoksi. Boreaaliset lehdot voivat ylläpitää myös monipuolista kääpälajistoa, mukaan lukien Punaisen listan lajistoa, mikäli lahopuuprofiilissa on riittävä määrä suurikokoista ja puulajistoltaan monipuolista lahopuuta. Lehtipuiden suuren osuuden vuoksi lehdot tarjoavat elinympäristöjä erityisesti lehtipuihin erikoistuneille kääville, ja siksi lehdoilla on täydentävä merkitys kääpien elinympäristönä boreaalisessa metsämaisemassa.
Kirjallisuus
Junninen K., Komonen A. (2011). Conservation ecology of boreal polypores: a review. Biological Conservation 144(1): 11–20. https://doi.org/10.1016/j.biocon.2010.07.010.
Kotiranta H., Junninen K., Saarenoksa R., Kinnunen J., Kytövuori I. (2010). Aphyllophorales & Heterobasidiomycetes. In: Rassi P., Hyvärinen E., Juslén A., Mannerkoski I. (eds). The 2010 Red List of Finnish species. Ministry of Environment & Finnish Environment Institute. Helsinki. s. 249–263. ISBN 978-952-11-3806-5. http://www.ymparisto.fi/punainenlista.
Nordén J., Penttilä R., Siitonen J., Tomppo E., Ovaskainen O. (2013). Specialist species of wood-inhabiting fungi struggle while generalists thrive in fragmented boreal forests. Journal of Ecology 101(3): 701–712. https://doi.org/10.1111/1365-2745.12085.
Siitonen J. (2001). Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forests as an example. Ecological Bulletins 49: 11–41.
Siitonen J., Martikainen P., Punttila P., Rauh J. (2000). Coarse woody debris and stand characteristics in mature managed and old-growth boreal mesic forests in southern Finland. Forest Ecology and Management 128(3): 211–225. https://doi.org/10.1016/S0378-1127(99)00148-6.