Matti Koivula  ,
Ilkka Vanha-Majamaa
,
Ilkka Vanha-Majamaa
Eri hakkuu- ja luonnonhoitomenetelmien vaikutukset monimuotoisuuteen Fennoskandiassa
Koivula M., Vanha-Majamaa I. (2021). Eri hakkuu- ja luonnonhoitomenetelmien vaikutukset monimuotoisuuteen Fennoskandiassa. Metsätieteen aikakauskirja vuosikerta 2021 artikkeli 10481. https://doi.org/10.14214/ma.10481
Tiivistelmä
Avohakkuisiin perustuvaa metsätaloutta on Fennoskandiassa harjoitettu kohta kiertoajan verran. Tämän seurauksena valtaosa Suomen metsistä on puustorakenteeltaan verrattain yksinkertaisia, vaihdellen alueellisella tasolla eri sukkessiovaiheiden kuviomosaiikkina. Samaan aikaan metsäpalot on tukahdutettu tehokkaasti. Toimien vuoksi sadat eliölajit ovat uhanalaistuneet. Uhanalaisen lajiston turvaamiseksi talousmetsien hoidossa tarvitaan parannuksia, jollaisiksi on ehdotettu säästöpuumetsätaloutta (variable retention forestry) ja erilaisia luonnonhoitotoimia. Tässä katsauksessa tarkastelemme kokeellisten tutkimuksien perusteella säästöpuumetsätalouden ja kahden luonnonhoitomenetelmän, kulotuksen ja lahopuun lisäämisen, monimuotoisuusvaikutuksia eri eliöryhmiin Fennoskandiassa. Tulosten perusteella (i) monet lajit reagoivat hakkuisiin lyhytaikaisesti positiivisesti johtuen vapautuneista resursseista, kuten ravinteista, valoisuuden lisääntymisestä, hakkuutähteistä ja kannoista; (ii) sulkeutuneiden metsien lajisto kärsii hakkuista, mutta säästettävä puusto edistää niiden säilymistä, ja jos puustosta jätetään 50–70 %, hakkuita edeltävä lajisto säilyy lähes ennallaan; (iii) hakkuiden aiheuttamat lajistomuutokset kestävät ainakin 10–30 vuotta, mutta esimerkiksi käävillä jopa sata vuotta; iv) kulotuksen välittömät vaikutukset ovat useimmissa eliöryhmissä negatiivisia, poikkeuksena kulonsuosijalajisto, mutta 10–15 vuoden jälkeen kulotusalojen lajistoon on jo palautunut harvinaisia ja uhanalaisia lajeja; (v) puiden päällä kasvavien epifyyttijäkälien palautuminen kulotuksen jälkeen kestää muuta lajistoa pidempään, vähintään 10–15 vuotta; (vi) lahopuun, kuten tekopökkelöiden, lisääminen suosii lahopuuta tarvitsevaa lajistoa, mutta tekopökkelöillä elävä lajisto poikkeaa luontaisesti kuolleiden puiden lajistosta; ja (vii) kosteusolojen ja metsikön sisäisen ympäristövaihtelun säilyminen hakkuiden yhteydessä on metsälajistolle tärkeää varsinkin kuusivaltaisissa metsissä. Hakkuumenetelmällä sinänsä on melko pieni merkitys uhanalaiselle lajistolle, mutta luonnonhoitotoimet ovat huomattavasti tärkeämpiä; hakkuissa jätettävä säästöpuusto vähentää hakkuiden negatiivisia lajistovaikutuksia sitä paremmin, mitä enemmän puustoa jätetään. Lajien säilymiselle on ensiarvoisen tärkeää kasvupaikkaolosuhteiden säilyminen ja jatkuvuus, lahopuun säästäminen ja tuottaminen, vanhojen puiden osuuden lisääminen ja sekapuustoisuuden ylläpitäminen.
Avainsanat
biodiversiteetti;
metsänhoito;
poimintahakkuu;
luonnonhoito;
pienaukkohakkuu;
säästöpuu;
tekopökkelö
-
Koivula,
Luonnonvarakeskus (Luke), Luonnonvarat, Helsinki
Sähköposti
matti.koivula@luke.fi
- Vanha-Majamaa, Luonnonvarakeskus (Luke), Luonnonvarat, Helsinki Sähköposti ilkka.vanha-majamaa@luke.fi
Vastaanotettu 17.11.2020 Hyväksytty 1.3.2021 Julkaistu 17.3.2021
Katselukerrat 148666
Saatavilla https://doi.org/10.14214/ma.10481 | Lataa PDF

Supplementary Files
 |
1 Johdanto
Yli 90 % Fennoskandian metsistä on talousmetsiä, joissa on jo kiertoajan verran harjoitettu intensiivistä metsätaloutta avohakkuineen ja maanmuokkauksineen. Tämä on johtanut valtaosin tasarakenteisiin ja -ikäisiin metsiköihin (Gustafsson ym. 2010) joissa lahopuuta on niukasti ja monimuotoisuuden kannalta tärkeää iäkästä puustoa on hyvin vähän. Avohakkuiden ekologisia vaikutuksia on tutkittu paljon viime vuosikymmeninä, ja niitä on vertailtu vaihtoehtoisten hakkuumenetelmien vaikutuksiin yhä enenevässä määrin (esim. Edenius ja Elmberg 1996; Koivula ym. 2014; Vanha-Majamaa ym. 2017).
Voimaperäisen metsätalouden rinnalla monimuotoisuuteen on vaikuttanut metsäpalojen tehokas sammutustoiminta. Metsäpalot eivät ole olleet suuressa roolissa Suomessa vuosikymmeniin tehokkaan palontorjunnan (metsäpalojen ennaltaehkäisy, havainnointi ja sammutus), tiheän metsätieverkon ja intensiivisen metsätalouden ansiosta. Tämä on vaikuttanut kuloja ja toisaalta myös kuollutta puuta vaativiin lajeihin negatiivisesti, mistä syystä kulotusta on alettu suositella luonnonhoidon keinona (mm. Kotiaho ym. 2015; Kontula ja Raunio 2018; Hyvärinen ym. 2019). Kulotusmäärät eivät suosituksista huolimatta ole kasvaneet, ja kulotuspinta-alat ovat Suomessa muutama sata hehtaaria vuodessa (Korhonen ym. 2017; Lindberg ym. 2020). Ruotsissa uudistusaloista FSC-sertifiointijärjestelmän mukaan vähintään 5 % pitää kulottaa. Vuosina 2011–2015 Ruotsin metsäpinta-alasta paloi luontaisesti tai kulotettiin keskimäärin vuosittain vain 0,006 % (Ramberg ym. 2018). Viime vuosina Ruotsissa on kuitenkin esiintynyt suurpaloja (Gustafsson ym. 2019): vuonna 2014 paloi noin 95 ja vuonna 2018 noin 210 neliökilometriä metsämaata (Larsson 2018).
Kuollut puu (alempana lahopuu) on ollut vuosikymmenet hyvin vähissä talousmetsissämme, mutta hiljattain sen säilyttämistä hakkuissa ja aktiivista lisäämistä on alettu suositella luonnonhoidon keinona (Tapio Oy 2019). Tästä huolimatta lahopuun määrä ei ole talousmetsissä kasvanut, ja Pohjois-Suomessa lahopuun määrä on jopa vähentynyt merkittävästi (Luonnonvarakeskus 2016). Jokainen koneellinen toimenpide metsissä tuhoaa varsinkin maalahopuustoa (Rabinowitsch-Jokinen ja Vanha-Majamaa 2010). Erityisen vahingollista lahopuun säilymiselle on uudistushakkuiden yhteydessä mahdollisesti tehtävä maanmuokkaus (Hautala ym. 2004). Lahopuun vähyys talousmetsissä onkin pääsyy 156 metsälajin uhanalaisuudelle (Hyvärinen ym. 2019). Tilanteen helpottamiseksi talousmetsiin on viime vuosikymmeninä alettu tehdä tekopökkelöitä katkaisemalla eläviä puita 3–5 m korkeudelta; suojelualueilla kuollutta puuta on tuotettu myös mm. kaulaamalla ja työntämällä puita nurin kaivinkoneella.
Vanhojen elävien ja kuolleiden puiden, vanhojen metsien niukkuuden ja metsäpalojen tehokkaan torjunnan vuoksi sadat metsälajit ovat nykyään uhanalaisia (Hyvärinen ym. 2019). Suomi on muiden EU-maiden tapaan sitoutunut monimuotoisuuden heikkenemisen pysäyttämiseen vuoteen 2030 mennessä (Euroopan unioni 2020). Monimuotoisuuden heikentymiseen vastataan luonnonhoitotoimilla ja suojelualueita perustamalla, mutta myös hakkuumenetelmien kirjo tarjoaa mahdollisuuksia yhdistää erilaisia tavoitteita. Talousmetsien hoitokäytäntöjä muuttamalla voidaan vaikuttaa huomattavasti suurempaan pinta-alaan kuin mihin suojelu- ja ennallistamistoimilla on mahdollista. Suomen metsien monimuotoisuuden ylläpitämisen ja palauttamisen kannalta talousmetsien käsittely onkin avainasemassa.
Maailmanlaajuisesti on otettu käyttöön säästöpuuhakkuumenetelmiä, joissa suuri osa puustosta jätetään hakkuissa uudistusalalle pystyyn, sekä ennallistamismenetelmiä, joilla uhanalaisille lajeille suotuisia olosuhteita voidaan palauttaa (Kuuluvainen ja Grenfell 2012; Beese ym. 2019). Suomalaisessa nykykäytännössä avohakkuulle jätetään useimmiten kymmenisen säästöpuuta hehtaarille yksittäin tai ryhmiin (enintään 2 % tilavuudesta; Kuuluvainen ym. 2019), mikä tulkitaan vielä avohakkuuksi (Fedrowitz ym. 2014). Säästöpuuhakkuulla viittaamme tässä artikkelissa suurempiin jätettäviin puumääriin, noin kymmenestä reiluun 50 prosenttiin puustosta, menetelmien ollessa poiminta- ja pienaukkohakkuiden muunnelmia. Näitä menetelmiä on tutkittu kokeellisesti enemmälti vasta 1990-luvulta lähtien. (Taulukko 1).
| Taulukko 1. Tiedossamme olevat pohjoismaiset säästöpuuhakkuu- ja luonnonhoitokokeet, joissa kustakin käsittelystä on vähintään kolme metsikkötason toistoa. Kokeiden nimikkeet eivät välttämättä ole virallisia; lisätiedot, katso nimikkeen perässä oleva numero ja vastaavat viitteet taulukon alareunassa. Perust = kokeen perustamisvuosi; Kesto = kokeen suunniteltu kesto (Pitkäaik = yli 10 vuotta); Metsät = valtapuulajit; Säästö = säästöpuita yksittäin tai (useimmiten) ryhminä avohakkuulla; Pien = pienaukkohakkuu (aukon läpimitta enintään 50 m); Poim = poimintahakkuu; Laho = lahopuun tuottaminen (useimmiten tekopökkelöt); Kulo = kulotus tehty ennallistamis-, luonnonhoito- tai metsänhoitotarkoituksessa; Muu = mahdolliset muut toistetut tekijät. Taulukon alaosassa tutkitut eliöryhmät ja muut tekijät seuraavasti: Kasvill = pohja- ja kenttäkerroskasvillisuus (putkilokasvit ja metsänpohjan sammalet ja jäkälät); Epif = epifyytti- ja/tai epiksyylisammalet ja jäkälät (erotettu osassa kokeita); Käävät = käävät; Maa = maan pinnassa elävät hyönteiset ja hämähäkit; Saproks = lahopuulla elävät (saproksyyli-) ja aktiivisesti lentävät kovakuoriaiset; Muu = mahdollisia muita eliöryhmiä (ks. Koivula ja Vanha-Majamaa 2020); Laho = järeä lahopuusto inventoitu; Puu = korjattu puusto mitattu; Sosiaal = esteettiset ja/tai sosiaaliset vaikutukset; Talous = hakkuutapojen taloudellinen vertailu. * = tutkimustuloksia ei tietääksemme vielä ole julkaistu vertaisarvioiduissa englanninkielisissä sarjoissa. | ||||||||||
| KOKEIDEN PERUSTIEDOT | ||||||||||
| Nimike | Maa | Perust | Kesto | Metsät | Säästö | Pien | Poim | Laho | Kulo | Muu |
| NaturKultur 1 | Ruotsi | 1989 | Pitkäaik | Seka | X | Istutus | ||||
| Härjedalen 2 | Ruotsi | 1989 | Pitkäaik | Mänty, kuusi | X | |||||
| Fagerön 3 | Ruotsi | 1994 | 5 vuotta | Seka | X | X | ||||
| MONTA 4 | Suomi | 1995 | Pitkäaik | Kuusi | X | X | X | X | ||
| Snöberget 5 | Ruotsi | 1997 | 5 vuotta | Kuusi | X | |||||
| RETREE 6 | Suomi | 1998 | Pitkäaik | Kuusi | X | X | Kosteus | |||
| FIRE 7 | Suomi | 2000 | Pitkäaik | Mänty | X | X | ||||
| Medelpad 8 | Ruotsi | 2000 | Ei tiedossa | Seka | X | |||||
| EVO 9 | Suomi | 2001 | Pitkäaik | Kuusi | X | X | X | Kosteus | ||
| Tammimetsä 10 | Ruotsi | 2001 | Pitkäaik | Tammi, seka | X | |||||
| Lahopuulisäys 11 | Suomi | 2002 | Pitkäaik | Mänty, kuusi | X | Kaato, kaulaus | ||||
| Leivonmäki 12 | Suomi | 2003 | Pitkäaik | Kuusi | X | Määrä, jakautum. | ||||
| Hedmark 13 | Norja | 2004 | 2 vuotta | Seka | Reunavaikutus | |||||
| PuroMONTA 14 | Suomi | 2004 | Pitkäaik | Kuusi | X | X | Puronvarret | |||
| Elimyssalo 15 | Suomi | 2005 | Pitkäaik | Mänty, kuusi | X | |||||
| UPM-koe 16 | Suomi | 2005 | 3 vuotta | Kuusi | X | X | ||||
| DISTDYN 17 | Suomi | 2009 | Pitkäaik | Mänty, kuusi | X | X | X | X | ||
| Eriksköp 18 | Ruotsi | 2009 | Pitkäaik | Mänty, kuusi | X | |||||
| Future Forest 19 | Ruotsi | 2010 | Pitkäaik | Mänty, kuusi | X | X | ||||
| Kuusilahopuu 20 | Ruotsi | 2011 | Ei tiedossa | Kuusi | X | |||||
| Rogberga 21 | Ruotsi | 2012 | Pitkäaik | Seka | X | X | ||||
| Suojuspuu 22 | Viro | 2012 | Pitkäaik | Mänty | X | Suojuspuu | ||||
| Effaråsen 23 | Ruotsi | 2012 | Pitkäaik | Mänty | X | X | X | |||
| UNEVEN 24 | Ruotsi | 2013 | Pitkäaik | Kuusi | X | |||||
| TUTKITUT LAJIRYHMÄT JA MUUT TEKIJÄT | ||||||||||
| Nimike | Kasvill | Epif | Käävät | Maa | Saproks | Muu | Laho | Puu | Sosiaal | Talous |
| NaturKultur | X* | X | X* | X* | ||||||
| Härjedalen | X | |||||||||
| Fagerön | X | |||||||||
| MONTA | X | X | X* | X | X | X | X | X | X | |
| Snöberget | X | |||||||||
| RETREE | X | X | X | X | X | X* | ||||
| FIRE | X | X | X | X | X | X | X | |||
| Medelpad | X | |||||||||
| EVO | X | X | X | X | X | X | X | X | ||
| Tammimetsä | X | X | X | X | X | X | ||||
| Lahopuulisäys | X | |||||||||
| Leivonmäki | X | |||||||||
| Hedmark | X | |||||||||
| PuroMONTA | X | X | X | X | X | |||||
| Elimyssalo | X | X | X | X | ||||||
| UPM-koe | X | X | X | |||||||
| DISTDYN | X | X | X | X | X | X* | X | X* | ||
| Eriksköp | X* | X | ||||||||
| Future Forest | X | X | X | X | X | X | ||||
| Kuusilahopuu | X | |||||||||
| Rogberga | X* | X* | ||||||||
| Suojuspuu | X | X | ||||||||
| Effaråsen | X | X | X | X | X | X | X | X | ||
| UNEVEN | X | X | X | |||||||
| 1 Djupström ja Weslien 2019; 2 Hagner 1992; 3 Lindhe ja Lindelöw 2004; 4 Kaila 1998; 5 Hedenås ja Ericson 2003; 6 Matveinen-Huju ym. 2006, Hautala ym. 2011; 7 Hyvärinen ym. 2005; 8 Perhans ym. 2009; 9 Toivanen ym. 2004, Lilja ym. 2007, Vanha-Majamaa ym. 2007; 10 Götmark ym. 2005; 11 Pasanen 2017; 12 Komonen ym. 2014, Elo ym. 2019, Norberg ym. 2019; 13 Fossestol ja Sverdrup-Thygeson 2009; 14 Toivanen ym. 2004, Selonen ja Kotiaho 2013; 15 Hekkala ym. 2014a, 2014b, 2016; 16 Rabinowitsch-Jokinen ja Vanha-Majamaa 2010; 17 Koivula ym. 2014; 18 Drössler 2016; 19 Hägglund ym. 2015; 20 Olsson ym. 2011; 21 Drössler 2016; 22 Tullus ym. 2019; 23 Djupström ja Weslien 2019; 24 Joelsson ym. 2017. | ||||||||||
Käsittelemme tässä artikkelissa säästöpuuhakkuiden ja luonnonhoidon vaikutuksia eri lajiryhmiin. Sisällytämme tarkasteluun myös hakkuisiin yleisesti kytkeytyvän maanmuokkauksen lajistovaikutukset. Talousmetsien luonnonhoidon menetelmistä keskitymme kulotuksiin ja kuolleen puun tuottamiseen. Aineistonamme on Fennoskandiassa (ml. Baltia) tehdyt kokeelliset säästöpuuhakkuu- ja luonnonhoitotutkimukset. Tarkoituksena on tarjota metsistä kiinnostuneelle lukijakunnalle näistä tutkimuksista kertynyt ekologinen tieto tiivistetyssä muodossa, joka on sisällytettävissä metsien käytön suunnitteluun ja toteutukseen.
2 Aineisto ja menetelmät
Tämän artikkelin pohjana on kirjallisuuskatsaus (Koivula ja Vanha-Majamaa 2020). Se pohjautui Northern European Database of Long-Term Forest Experiments -tietokantaan (noltfox.metla.fi) ja Web of Science -tietokannasta 30.9.2019 tekemäämme artikkelihakuun, jossa käytimme seuraavaa termiketjua: (forest* AND (Finland OR Sweden OR Norway OR Estonia OR Latvia OR Lithuania) AND (logg* OR fell* OR harvest* OR cut* OR select* OR thinn* OR gap OR deadwood OR dead wood OR fire OR burn* OR restor* OR stump*) AND (plant* OR moss* OR lichen* OR polypor* OR fung* OR beetle* OR insect* OR diversit*). Mainittuun katsaukseen valitsimme ne artikkelit, joiden tulokset olivat peräisin sellaisista empiirisistä kokeista, joissa kutakin vertailtavaa käsittelyä edusti vähintään kolme metsikköä. Täydensimme hakua artikkelien ensimmäisen ja senioritekijän nimillä sekä artikkelien lähdeluetteloiden avulla. Käsillä olevaa katsausta varten täydensimme aineistoa kahdella ennallistamiskokeella (Leivonmäki ja Vienansalo; Taulukko 1). Tämä katsaus siis perustuu kaikkiaan 24 luonnonhoito- tai hakkuukoeasetelmaan, joista on monimuotoisuutta käsitteleviä, vertaisarvioituja julkaisuja (Kuvat 1–2). Pääosa tuloksista on saatu enintään 10–15 vuotta hakkuista (Luku 3, Tulokset), joten niiden tarkastelussa (Luku 4, Tulosten tarkastelu) tukeudumme lisäksi kronosekvenssitutkimuksiin (eri aikoina käsiteltyjen kohteiden vertailu tiettynä ajanhetkenä), joilla päästään kiinni pitempiaikaisiin muutoksiin.
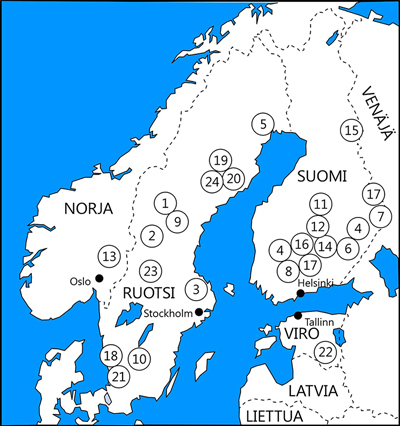
Kuva 1. Tarkasteltujen pohjoismaisten säästöpuu- ja ennallistamiskokeiden sijainnit (vertaa Taulukko 1). Numerot viittaavat kokeisiin seuraavasti: 1 NaturKultur; 2 Härjedalen; 3 Fagerön; 4 MONTA (läntinen ja itäinen alue; numero kartalla kahdesti); 5 Snöberget; 6 RETREE; 7 FIRE; 8 EVO; 9 Medelpad; 10 Tammimetsä; 11 Lahopuulisäys (30 metsikköä tasaisesti Etelä- ja Keski-Suomessa); 12 Leivonmäki; 13 Hedmark; 14 PuroMONTA; 15 Elimyssalo; 16 UPM-koe; 17 DISTDYN (läntinen ja itäinen alue; numero kartalla kahdesti); 18 Eriksköp; 19 Future Forest; 20 Kuusilahopuu; 21 Rogberga; 22 Suojuspuu; 23 Effaråsen; 24 UNEVEN. Symboleja on siirretty satunnaisiin suuntiin päällekkäisyyden välttämiseksi.

Kuva 2. Käytännön säästöpuu- ja ennallistamisesimerkkejä. Ylh. vas.: säästöpuuryhmiä avohakkuulla, ensimmäinen hakkuun jälkeinen kesä (MONTA, kesäkuu 1996). Ylh.oik.: poimintahakattu eri-ikäisrakenteinen sekametsä, jossa koivutekopökkelö, toinen hakkuun jälkeinen kesä (DISTDYN, kesäkuu 2012). Alh.vas.: tekopökkelöitä avohakkuulla, toinen hakkuun jälkeinen kesä (DISTDYN, kesäkuu 2012). Alh.oik.: ennallistamispoltto, johon yhdistetty kaatamalla tuotettua lahopuuta korpipainanteessa ja sen lähellä, seitsemäs hakkuun jälkeinen kesä (EVO, toukokuu 2009). Kuvat: Matti Koivula.
Perustelemme keskittymistä kokeelliseen tutkimukseen seuraavasti. Ensiksikin lajisto usein tunnetaan ennen toimenpiteitä. Toiseksi toimenpiteet tehdään samalla tavoin useissa paikoissa, millä päästään kiinni vaikutuksien vaihteluun. Kolmanneksi kokeissa on verrokkimetsiköitä, jotka ovat muuten samanlaisia kuin toimenpidemetsiköt, mutta niissä ei tehdä toimenpiteitä. Nämä seikat lisäävät ratkaisevasti tulosten luotettavuutta.
Tämän narratiivimuotoisen artikkelin pohjana olleessa artikkelissa (Koivula ja Vanha-Majamaa 2020) lajiston vasteita tarkasteltiin lajiryhmittäin. Tässä artikkelissa tarkastelemme vasteita toimenpide kerrallaan, raportoiden eri hoitotoimenpiteiden positiiviset, neutraalit ja negatiiviset vaikutukset. Painotamme eliöryhmiä, joissa on paljon elinympäristönsä tai kasvualustansa suhteen erikoistuneita tai uhanalaisia lajeja. Perustelemme painotusta sillä, että monimuotoisuuden säilymisen kannalta juuri nämä lajit vaativat erityistoimia, eivät niinkään runsastuvat tai lähes kaikkialla tavattavat yleislajit. Kattava lista tarkastelujen pohjana olevista artikkeleista löytyy liitetiedostosta L1.
3 Tulokset
3.1 Säästöpuuhakkuiden lajistovaikutukset
Hakkuuintensiteetin nosto kasvattaa useiden eliöryhmien kokonaislajimäärää metsikkötasolla, etupäässä siten, että yleis- ja avomaalajit runsastuvat ja sulkeutunutta latvuskerrosta vaativat lajit vähenevät (Taulukko 2). Pienet säästöpuuhakkuussa jätetyt puuryhmät (23–50 m3 ha–1 2–3 ryhmänä) muuttuvat lajikoostumukseltaan voimakkaasti, joskin monet tavalliset sulkeutuneen metsän lajit säilyvät niissä ainakin kymmenen vuotta (Siira-Pietikäinen ja Haimi 2009; Johnson ym. 2014; Vanha-Majamaa ym. 2017; Koivula ym. 2019). Sitä vastoin sellaiset poiminta- tai pienaukkohakkuut, joissa puustosta säästetään vähintään 50 %, eivät näytä juurikaan muuttavan metsikön kasvi- ja eläinlajistoa (Koivula 2002; Pietikäinen ym. 2003; Matveinen-Huju ja Koivula 2008; Joelsson ym. 2018). Esimerkiksi haavan (Populus tremula L.) epifyyttijäkälät näyttävät säilyvän, joskin niiden kasvu hidastuu (Hedenås ja Ericson 2003).
| Taulukko 2. Yhteenveto taulukon 1 kokeiden lajiryhmäkohtaisista julkaistuista havainnoista. Lyhyen (enintään kolme vuotta toimenpiteistä) ja pitkän aikavälin vaikutukset erotettu vinoviivoilla. + = positiivinen, - = negatiivinen ja ± = ei havaittua tai epäselvä vaikutus, ja na = ei tutkittu tai ei julkaistu. Säästö = hakkuussa säästetty elävä puusto; Kulot = metsän polttaminen; Lahop = järeän lahopuun tuottaminen; Kosteat = koeasetelmaan sisällytetty kosteat ja märät pienympäristöt, kuten korvet; Muokk = maanmuokkaus; Vuorov = em. tekijöiden merkittävät vuorovaikutukset; Muut tekijät = muut raportoidut lajimäärään tai runsauksiin vaikuttavat tekijät. 1 = lajiryhmässä on uhanalaista tai pitkälle erikoistunutta lajistoa. | |||||||
| Lajiryhmä | Säästö | Kulot | Lahop | Kosteat | Muokk | Vuorov | Muut tekijät |
| Aluskasvillisuus | |||||||
| Myöhäissukkession lajit | + / + | - / - | na / na | + / + | - / - | ||
| Pioneerilajit | - / - | + / + | na / na | na / na | + / + | ||
| Sammalet ja jäkälät | + / + | - / na | na / na | + / + | - / - | ||
| Epifyytti- ja epiksyylisammalet ja -jäkälät | |||||||
| Epifyytit 1 | + / + | - / - | na / na | + / na | - / na | Kulotuksen pitkäaikaisvaikutus ehkä positiivinen | |
| Epiksyylit 1 | + / + | - / na | na / na | na / na | - / - | ||
| Sienet | |||||||
| Käävät 1 | ± / + | - / + | + / + | ± / ± | na / na | Säästö × Kulot | Kuollut maapuu tekopökkelöä parempi |
| Lakkisienet | na / na | na / + | na / na | na / na | na / na | ||
| Maan pinnan niveljalkaiset | |||||||
| Maakiitäjäiset, varjolajisto | + / + | - / - | na / + | na / ± | - / na | Metsikön sisäinen vaihtelu usein hakkuutapaa merkittävämpi | |
| Maakiitäjäiset, avomaalajisto | - / - | + / + | na / - | na / - | + / na | Metsikön sisäinen vaihtelu usein hakkuutapaa merkittävämpi | |
| Maakiitäjäiset, varjostusgeneralistit | - / - | - / + | na / ± | na / - | ± / na | ||
| Maakiitäjäiset, korpilajit | + / + | na / - | na / ± | + / + | na / na | ||
| Hämähäkit, metsän verkonkutojat | + / na | na / na | na / na | na / na | na / na | ||
| Hämähäkit, juoksuhämähäkit | - / na | na / na | na / na | na / na | na / na | ||
| Lukit | + / na | na / na | na / na | na / na | na / na | ||
| Lahopuu- ja muut aktiivisesti lentävät kovakuoriaiset | |||||||
| Tuoreen kuolleen puun lajit 1 | + / ± | + / ± | + / ± | na / na | na / na | Kulot × Lahop | Lahopuun laatu tärkeä; jos lahopuuta paljon, hakkuutavan merkitys pieni |
| Pitkälle lahonneen puun lajit 1 | ± / + | ± / + | ± / + | na / na | na / na | Kulot × Lahop | Lahopuun laatu tärkeä; jos lahopuuta paljon, hakkuutavan merkitys pieni |
| Sienensyöjät 1 | + / + | - / - | + / + | na / na | na / na | Kulot × Lahop | Lahopuun laatu tärkeä; jos lahopuuta paljon, hakkuutavan merkitys pieni |
| Muut kuin lahopuulajit | - / - | + / + | ± / ± | na / na | na / na | ||
| Muut eliöryhmät | |||||||
| Latikat 1 | ± / + | + / + | na / + | na / na | na / na | ||
| Kekomuurahaiset | ± / na | na / - | na / ± | na / na | na / na | ||
| Juoksujalkaiset | + / + | na / na | na / na | na / na | na / na | ||
| Hyppyhäntäiset ja punkit | + / + | na / na | na / na | na / na | na / na | ||
| Maaperän hajottajat | + / + | na / na | na / na | na / na | - / - | ||
| Pölyttäjähyönteiset | na / - | na / + | na / ± | na / na | na / na | ||
| Loiskärpäset | na / - | na / ± | na / ± | na / na | na / na | ||
| Sienisääsket | + / na | na / na | + / na | na / na | na / na | ||
| Vaivaispäästäinen | + / na | na / na | na / na | na / na | na / na | ||
| Metsäsopuli | + / na | na / na | na / na | na / na | na / na | ||
| Pohjanlepakko | na / + | na / na | na / na | na / na | na / na | ||
Vanha-Majamaa ym. (2017) havaitsivat, että 10 vuoden kuluttua hakkuista sammallajeja oli hävinnyt enemmän 67 % säästäneessä poiminta- kuin 50 % säästäneessä pienaukkohakkuussa, mikä viittaa siihen, että säästöpuuston keskittäminen ryhmiin säilyttää tätä lajistoa paremmin, kuin jos puita säästetään tasaisesti. Toisaalta puolen hehtaarin säästöpuuryhmät – läpimitaltaan 50–60 m – eivät kykene säilyttämään herkimpiä maksasammalia tai epiksyyli- ja epifyyttijäkäliä (Perhans ym. 2009; Hautala ym. 2011; Hämäläinen ym. 2016). Vastaavasti puronvarsien kasvilajisto säilyy, jos hakkaamaton suojavyöhyke on 35–45 m (Oldén ym. 2019a). Havainnot viittaavat siihen, että pienilmasto- ja valaistusolot (Oldén ym. 2019b), ja näiden myötä lajisto, muuttuvat avohakkuun seurauksena vähintään 25–30 metrin etäisyydelle – varttuneissa eteläsuomalaisissa metsissä siis karkeasti valtapuiden pituuden verran – säästöpuuryhmän tai reunametsän sisäosiin.
Hakkuut vaikuttavat haitallisesti vaateliaaseen metsälajistoon, mutta säästöpuuston jättäminen vähentää vaikutuksia (Taulukko 2). Hakkuutähteet (oksat, latvukset ja kannot) hyödyttävät monia kääpiä ja kovakuoriaisia, myös uhanalaisia lajeja. Näiden lajiryhmien kokonaisyksilö- ja lajimäärät kuitenkin palautuvat lähtötasolle muutamassa vuodessa, ja pääosin hyötyjät lienevät tuoretta lahopuuta vaativia lajeja (Hyvärinen ym. 2006; Toivanen ja Kotiaho 2007a, 2007b; Heikkala ym. 2016a, 2016b; Suominen ym. 2018; Jokela ym. 2019). Jos hakkuualalle on jätetty elävää säästöpuustoa 10 m3 ha–1, määrän nosto esimerkiksi 50 m3 ha–1:in vaikuttaa lahopuuta tarvitseviin kääpiin ja kovakuoriaisiin lyhyellä aikavälillä varsin vähän (Hyvärinen ym. 2005, 2009; Toivanen ja Kotiaho 2007a, 2007b, 2010; Heikkala ym. 2016a). Säästöpuiden hiljalleen kuollessa voisi olettaa niiden määrän kasvattamisella olevan positiivinen pitkän aikavälin vaikutus tähän lajistoon; tämä asia kuitenkin vaatii lisätutkimuksia.
3.2 Kulotuksen lajistovaikutukset
Kulotuksen välittömät lajistovaikutukset ovat pääsääntöisesti haitallisia (Vanha-Majamaa ym. 2007; Berglund ym. 2011; Rudolphi ym. 2011; Hekkala ym. 2014a) kuloja vaativia tai suosivia lajeja lukuun ottamatta (Johansson ym. 2010; Hägglund ym. 2015; Heikkala ym. 2017). Myöhemmin paloalueet hyödyttävät jo suurta joukkoa harvinaisia ja uhanalaisia lajeja (Hyvärinen ym. 2006; Toivanen ja Kotiaho 2007a, 2007b; Junninen ym. 2008; Suominen ym. 2018). Kuitenkin puiden rungoilla elävät epifyyttijäkälät kärsivät kulotuksesta ainakin 10–15 vuotta: tuli tappaa yksilöitä suoraan ja ainakin tilapäisesti heikentää niiden kasvualustan laatua ja pienilmasto-oloja (Hämäläinen ym. 2014).
Kulotus muuttaa nopeasti lahopuuta vaativan kovakuoriaisyhteisön koostumuksen ja rakenteen, mikä saattaa samalla muuttaa niiden toiminnallista roolia ekosysteemissä (Heikkala ym. 2016b; Hägglund ja Hjältén 2018). Monimuotoisuushyödyt voivat kasvaa palaneen puun määrän kasvun myötä, kuten lahopuukuoriaisilla (Hekkala ym. 2014b; Komonen ym. 2014) ja käävillä (Suominen ym. 2015), tai säilyä ennallaan, kuten luteisiin kuuluvilla latikoilla (Aradidae) (Heikkala ym. 2017). Lajistovaikutuksen voimakkuus riippuu palaneen puun laadusta (käävät; Suominen ym. 2019), maan pinnan rakenteista (pölyttäjähyönteiset; Rodriguez ja Kouki 2017) ja maaperän kosteudesta (korpien kovakuoriaiset; Toivanen ym. 2014).
3.3 Lahopuun tuottamisen lajistovaikutukset
Tekopökkelöt ja muut tuotetut lahopuut hyödyttävät useita lahopuulla eläviä lajiryhmiä, joissa on paljon harvinaista ja uhanalaista lajistoa, kuten kääpiä ja kovakuoriaisia (Toivanen ja Kotiaho 2007a, 2007b; Olsson ym. 2011; Pasanen ym. 2014). Erityisesti lahopuun lisäyksestä hyötyvät nilaa ja puuainesta syövät kovakuoriaiset (Toivanen ja Kotiaho 2010). Luonnollisesti kuolleisiin puihin verrattuna tuotetun lahopuun kääpälajisto on niukempaa ja koostumukseltaan osin erilaista (Pasanen ym. 2018, 2019).
Lahopuun tuottamisen lajistohyödyt riippuvat puulajista (Lindhe ja Lindelöw 2004; Toivanen ja Kotiaho 2010) ja tuottamistavasta (Hägglund ja Hjältén 2018), mutta mikään puulaji ei uhanalaisen lajiston kannalta ole hyödytön. Tuotetuilla maapuilla elää enemmän kääpälajeja kuin tekopökkelöillä (Lindhe ym. 2004). Toisaalta tekopökkelöillä elää maapuita enemmän jälttä ja sienirihmastoa syöviä sekä uhanalaisia kovakuoriaislajeja (Gibb ym. 2006; Johansson ym. 2006; Hjältén ym. 2010; Andersson ym. 2015). Erityisesti myöhäisen lahoamisvaiheen puuta vaativat lajit hyötyvät läpimitaltaan suuremmista tekopökkelöistä (Jonsell ym. 2004; Lindhe ym. 2004, 2005). Koska tuotetun lahopuun lahoamisen edetessä siinä elävä lajisto muuttuu (Lindhe ja Lindelöw 2004), ei lyhyen aikavälin seuranta kovin hyvin kuvaa lajistolle koituvaa kokonaishyötyä. Toisaalta lahopuun sijainti suhteessa metsänreunaan vaikuttaa mm. puuaineksen kuivumiseen ja sitä kautta siinä elävään lajistoon (Fossestol ja Sverdrup-Thygeson 2009), millä voi olla merkitystä tekopökkelöiden tehokkaan sijoittelun kannalta; tätä reunavaikutusta olisi kuitenkin vielä tutkittava enemmän.
3.4 Hakkuiden, ennallistamisen ja metsärakenteiden yhteisvaikutukset
Jätettävä puusto kulotuksen yhteydessä (ja muutoinkin) ylläpitää varjostusta ja lisää seuraavan puusukupolven rakenteellista vaihtelua sekä ylläpitää jossakin määrin hakkuita edeltävää pienilmastoa. Tästä syystä säästöpuusto auttaa säilyttämään kuloalan polttoa edeltävää aluskasvillisuutta ja nopeuttaa sen palautumista (Heikkala ym. 2014; Johnson ym. 2014). Puusto myös suojaa maan pintakerrosta paahteelta ja parantaa siten varpujen marjasatoja (Granath ym. 2018) ja säilyttää kekomuurahaisten (suku Formica) pesäkumpuja (Toivanen ym. 2014). Kulotuksen negatiivinen vaikutus kääpiin lievenee jätettäessä säästöpuita (Suominen ym. 2015), ja monet harvinaiset tai uhanalaiset kovakuoriaislajit hyötyvät säästöpuista (Komonen ym. 2014).
Hakattavassa tai poltettavassa metsikössä olevat märät räme- tai korpipainanteet ovat tärkeitä näihin ympäristöihin erikoistuneille lajeille. Osa näistä lajeista voi selviytyä hakatulla tai poltetulla paikalla useita vuosia, kuten korpiympäristöissä elävä liekokurekiitäjäinen (Platynus mannerheimii Dejean) (Toivanen ym. 2014). On mahdollista, että puuston haihduttavan vaikutuksen poistuessa vedenpinnan kohoaminen suosii kosteita painanteita vaativia lajeja. Toisaalta poltetulle alalle jätetyt puuryhmät auttavat sulkeutuneen metsän maakiitäjäislajistoa (Martikainen ym. 2006a). Nämä havainnot viittaavat käsiteltävän metsikön rakenteellisen monipuolisuuden olevan tärkeä lajistoa säilyttävä piirre erityisesti häiriötilanteissa, kuten hakkuun tai metsäpalon yhteydessä.
Tässä artikkelissa käsitellyissä kokeissa on hakkuun yhteydessä usein tehty maanmuokkaus, mikä on tavanomainen toimi metsien uudistamisen yhteydessä. Maanmuokkaus vaikuttaa useimpiin sulkeutuneen metsän oloja vaativiin kasvi- ja hyönteislajeihin negatiivisesti (Hautala ym. 2011; Vanha-Majamaa ym. 2017; Tullus ym. 2018). Metsän tavalliseen kasvi- ja maakiitäjäislajistoon toimenpide vaikuttaa etupäässä metsikön sisäisesti siten, että yksilöt jakautuvat muokattuihin (avoimen ympäristön lajit, pioneerilajit) ja muokkaamattomiin maastonkohtiin (varjoisuutta vaativat lajit). Tämä uusjako voi näissä lajiryhmissä vaikuttaa maastonkohdassa havaittavaan yksilömäärään enemmän kuin itse hakkuu (Koivula 2002; Pihlaja ym. 2006; Toivanen ym. 2014; Vanha-Majamaa ym. 2017). Monimuotoisuuden säilymisen kannalta on merkittävää, että äestystä huomattavasti kevyempi laikkumätästyskin vähentää sekä epiksyylisammalien ja -jäkälien lajimäärää että näiden lajien kasvualustakseen vaatiman, pitkälle lahonneen puun määrää jopa alle puoleen (Rabinowitsch-Jokinen ja Vanha-Majamaa 2010; Hautala ym. 2011; Rabinowitsch-Jokinen ym. 2012).
4 Tulosten tarkastelu
Tässä osiossa tarkastelemme edellä kerrottuja tuloksia muuhun pohjoismaiseen metsäekologiseen tutkimukseen kytkettyinä. Tarkoituksenamme on ulottaa tarkastelu pitemmille ajanjaksoille kuin kokeellisten tutkimuksien avulla toistaiseksi voidaan tehdä. Tarkastelun pohjalta pyrimme tarjoamaan ehdotuksia metsien hoidon kehittämiseksi sekä osoittamaan tutkimustiedon aukkoja tulevan tutkimuksen tehokkaammaksi suuntaamiseksi.
4.1 Säästöpuiden merkitys hakkuissa
Säästöpuuston määrän ja puuryhmien koon kasvattaminen vähentävät hakkuiden negatiivisia lajistovaikutuksia. Yleisellä tasolla harvinaiset ja uhanalaiset lajit hyötyvät korkeammasta säästöpuuston määrästä; sama havainto on tehty harvennushakatuissa pohjoissuomalaisissa metsissä (Sippola ym. 2004). Metsikkötasolla yli 50 %:n säästöpuusto tai yli puolen hehtaarin säästöpuuryhmät tukevat sulkeutuneen metsän lajistoa jo varsin hyvin. Mallitustutkimuksissa on havaittu tämän tyyppisen jatkuvapeitteisen metsänkäsittelyn ylläpitävän kuusikoissa hyviä mustikkasatoja (Pukkala ym. 2011; Miina ym. 2020). Puuryhmien voi myös ajatella suojaavan monia monimuotoisuuden säilymisen kannalta välttämättömiä metsien rakennepiirteitä, kuten pökkelöitä, kuollutta maalahopuuta ja korpipainanteita, ja ne myös lisäävät uudistuvan metsän rakenteellista vaihtelua (Kruys ym. 2013). Sellaisenaan hyvin pienet, esimerkiksi noin kymmenen puun, säästöpuuryhmät eivät turvaa uhanalaista metsälajistoa mitenkään, ellei hakkuualueiden lahopuuston säilymiseen ja säästöpuuston järeyteen kiinnitetä hakkuukohteissa erityistä huomiota.
Avohakkuiden vaikutukset näkyvät kasvillisuudessa useita vuosikymmeniä esim. sammalilla, maksasammalilla ja varvuilla (Vanha-Majamaa ja Reinikainen 2000; Hedwall ym. 2013; Tonteri ym. 2016; Vanha-Majamaa ym. 2017). Toisaalta kuusivaltaisten ruotsalaismetsien kääpälajistossa näkyi satakunta vuotta aiemmin tehtyjen poimintahakkuiden vaikutus (Bader ym. 1995). Hakkuissa syntyneiden kantojen pohjapinta-alan kasvu (1,25–93,9 m2 ha–1) siis näkyi edelleen kääpien kokonaislajimäärän ja kantotiheyden kasvu (29–465 kpl ha–1) lisäksi uhanalaisten kääpien lajimäärän merkitsevänä laskuna. Havainnot viittaavat siihen, että kääpälajiston täydellinen palautuminen intensiivisen hakkuun jälkeen vaatii vanhan metsän rakenteiden ja pienilmaston palautumista, eli ainakin kiertoajan verran.
Metsikkötasolla säästöpuita voidaan jättää tasaisesti, ryhmittäin tai näiden yhdistelmänä. Vertailuja näiden tapojen välillä on tehty suhteellisen vähän monimuotoisuusnäkökulmasta. Pohjois-Suomen kuusikoissa pysty- ja maapuiden pintajäkälien (epifyytti- ja epiksyylilajit) lajimäärä oli suurempi puuryhmissä kuin yksittäin säästetyillä puilla (Ylisirniö ja Hallikainen 2018). Tulos on samanlainen kuin Vanha-Majamaan ym. (2017) havainto poiminta- ja pienaukkohakkuun vaikutuksista sammalten lajimäärään (katso Tulokset, kohta 3.1). Säilyttääkseen sulkeutuneen metsän pienilmaston olisi avohakkuille jätettävien puuryhmien oltava reilusti yli puolen hehtaarin kokoisia (katso Tulokset, kohta 3.1). Näiden tulosten mukaan puiden säästäminen ryhminä toimii tasaisesti jättämistä paremmin sulkeutuneen metsän lajiston kannalta.
Isot säästöpuut hyödyttävät järeää lahopuuta vaativaa lajistoa, sillä ne takaavat uuden lahopuun syntymisen jopa vuosikymmeniä, kun säästöpuita vähitellen kaatuu ja kuolee (Junninen ym. 2008; Berglund ym. 2011; Suominen ym. 2015; Hämäläinen ym. 2016). Monet harvinaiset ja uhanalaiset käävät ja jäkälät vaativat kasvualustoikseen erittäin järeitä puita ja ne ovat usein myös hitaita levittäytymään, joten isojen puiden pysyvä säästäminen on niiden säilymisen kannalta välttämätöntä (Hämäläinen ym. 2014; Suominen ym. 2018, 2019).
Yhdenkään uhanalaisen lajimme ei tiedetä tarvitsevan uudistushakkuita, vaikkakin monet menestyvät uudistusaloille jätetyillä suurilla elävillä ja kuolleilla puilla (Kaila ym. 1997; Martikainen 2001; Junninen ym. 2007), mutta jotkin erityishakkuut voivat joissakin tilanteissa olla lajiston suojelun kannalta perusteltuja. Esimerkiksi varovaiset harvennushakkuut voivat hyödyttää harvinaisia jäkälälajeja (Storaunet ym. 2008, 2014; Lõhmus ym. 2012). Ruotsissa tammivaltaisia metsiä on hoidettu muihin kuin tammeen (Quercus robur L.) kohdistuvin poimintahakkuin, mikä on hyödyttänyt uhanalaista tammella elävää lajistoa (Paltto ym. 2008; Nordén ym. 2012). Myös Suomessa lehtometsien luonnonhoitoon kuuluu alikasvoskuusten (Picea abies (L.) H. Karst.) poisto (Saaristo ja Vanhatalo 2019).
4.2 Kulotuksen ja lahopuun tuottamisen vaikutukset
Kulotus hyödyttää kulonvaatija-, paahdeympäristö- ja avoimen kasvupaikan lajistoa (Taulukko 2; Wikars 1995). Hyötyjiin kuuluvat myös monet lahopuulajit kulotuksessa syntyvän runsaan kuolevan ja kuolleen puuston ansiosta. Toisaalta kulotus voi tuhota 40 % lähtötilanteen kuolleesta puustosta ja vähentää kenttäkerroskasvillisuuden peittävyyttä 60–98 % (Eriksson ym. 2013). Epifyyttijäkälät kärsivät kulotuksesta ainakin 10–15 vuotta (Hämäläinen ym. 2014). Kulotukset olisi keskitettävä metsätyyppeihin, jotka luontaisestikin palavat useasti, kuten harju- ja paahdeympäristöihin ja mäntykankaisiin.
Lahopuututkimus on paljolti korostanut lahopuun määrää, mutta myös laatu ja monipuolisuus ovat tärkeitä uhanalaisen lajiston kannalta. Monet uhanalaiset lajit ovat erikoistuneet tiettyyn puulajiin, järeyteen ja lahoasteeseen (Junninen ja Komonen 2011). Pysty- ja maapuut ylläpitävät erilaista lajistoa, ensin mainittujen ollessa erityisen hyödyllisiä uhanalaiselle kovakuoriaislajistolle (Taulukko 2; Sverdrup-Thygeson ja Ims 2002; Andersson ym. 2015), jälkimmäisten taas kääpälajistolle (Junninen ja Komonen 2011). Tekopökkelöiden ja yleisesti lahopuun lajisto muuttuu lahoamisen edetessä (Lindhe ja Lindelöw 2004; Lindbladh ym. 2007; Jonsell ja Hansson 2011; Jonsell ym. 2019; Sandström ym. 2019). Lahoamisprosessia määräävä lahottajasienilajisto vaikuttaa tekopökkelöiden kovakuoriaislajistoon esimerkiksi siten, että sieni vaikuttaa lahoamisnopeuteen ja sitä kautta siihen, kuinka kauan tietyt kuoriaislajit voivat puuta asuttaa (Jonsell ym. 2005; Abrahamsson ym. 2008).
Metsätyyppi ja säästöpuuryhmän koko vaikuttavat sekä suoraan että epäsuorasti eliölajistoon. Epäsuorasti siksi, että ne vaikuttavat mm. paikan lahopuujatkumoon. Tavanomaisessa mustikkatyypin (MT) kuusikossa on usein pienialaisia ja siten usein kuviotiedoista puuttuvia korpipainanteita, joissa voi olla selvästi enemmän lahopuuta kuin niitä ympäröivissä kangasmetsissä (esim. 16 ja 2 m3 ha–1; Vanha-Majamaa ja Jalonen 2001). Näin ollen korpipainanteiden säästämisellä voidaan parhaassa tapauksessa säästää pääosa metsikön lahopuustosta (Hautala ym. 2004). On kuitenkin syytä huomata, että avohakkuun ympäröimien korpipainanteiden puusto on herkempää tuulenkaadoille kuin kangasmetsien (Hautala ja Vanha-Majamaa 2006; Hautala ym. 2011). Jos tavoite on lisätä lahopuun määrää nopeasti, säästöpuiden keskittäminen korpipainanteisiin voi tuottaa tulosta suhteellisen pian. Myös mäntykankailla pääosa 10 m3 ha–1 säästöpuustosta voi kaatua jo muutamassa vuodessa (Heikkala ym. 2014), eikä säästöpuusto näin ollen juuri ylläpidä pitkäaikaista lahopuujatkumoa. Jos säästöpuustoa jätetään enemmän kuin 10 m3 ha–1, sitä kaatuu hitaammin (Heikkala ym. 2014). Mäntyvaltaisissa metsissä säästöpuusto säilyy paremmin myös silloin, kun puut ovat pienempiä, ryhmissä ja käsittävät enemmän mäntyä (Pinus sylvestris L.) ja koivuja (Betula spp.) kuin kuusta (Hämäläinen ym. 2016). Näin ollen korpilaikun säästäminen sen rajaa myöten tai vähäinen järeä säästöpuusto kangasmaalla voivat tuottaa uutta lahopuuta nopeasti mutta lyhytkestoisesti, kun taas jatkumon kannalta suuremmat säästöpuumäärät ryhmissä ja kangasmailla toimivat paremmin. Lisäksi on syytä huomata, että topografialla on suuri merkitys säästöpuiden säilymiseen. Säästöpuiden jättäminen maastonkohtiin, jotka ovat paremmin suojassa vallitsevilta tuulilta, parantaa olennaisesti puiden säilymistä pystyssä.
Lahopuun tuottamisen puita tappamalla tai vahingoittamalla voisi kuvitella lisäävän kaarnakuoriaisepidemian tai metsäpalon riskiä. Kuitenkin mänty- ja kuusimetsissä tehtyjen hakkuu- ja ennallistamiskokeiden perusteella kaarnakuoriaisriski on pieni (Eriksson ym. 2006; Martikainen ym. 2006b; Toivanen ym. 2009). Kulotus voi vähentää tätä riskiä entisestään (Toivanen ym. 2010). Kuitenkin Kärvemo ym. (2017) havaitsivat Ruotsissa sekametsän kulotuksen tilapäisesti nostaneen sellaisten kaarnakuoriaisten (Curculionidae: Scolytinae) määriä, jotka kykenevät asuttamaan eläviä puita, kuten kirjanpainaja (Ips typographus L.) ja kuusentähtikirjaaja (Pityogenes chalcographus L.). Kirjoittajat arvelivat tämän johtuneen siitä, että kokeessa tehty kulotus ei yleisesti ottaen riittänyt tappamaan vaan ainoastaan heikentämään puita, jollaisiin nämä kaarnakuoriaiset nopeasti hakeutuvat. Lahopuun lisäämis- ja ennallistamisprojekteissa olisikin vältettävä etenkin kuusilahopuun tuottamista puita vahingoittamalla.
Hiljattain on ennustettu boreaalisten metsien kohtaavan lähivuosikymmeninä entistä useammin – tai poikkeavasti ajoittuen – myrsky- ja kuivuusjaksoja (Seidl ym. 2017). Tällöin saattavat hyönteisepidemioiden ohella myös metsäpalot yleistyä (Lehtonen ja Venäläinen 2020; Lindberg ym. 2020). Ruotsissa on viime vuosina esiintynyt suurpaloja: vuosina 2014 ja 2018 paloi satoja neliökilometrejä pääasiassa vaihtelevan ikäisiä, mäntyvaltaisia talousmetsiä (mm. Gustafsson ym. 2019). Näistä paloalueista jopa 60 % on suojeltu ja tutkimuskäytössä. Suomessa metsäpaloalueita ei juuri suojella johtuen lähinnä metsänomistajille tarjottujen korvauksien pienuudesta. Suojelemalla metsäpaloalueita voitaisiin kuitenkin säästää ennallistamispolttojen ja palaneiden puiden korjuun kustannuksissa ja parantaa samalla monien uhanalaisten lajien säilymistä.
4.3 Mihin tutkimusta pitäisi suunnata?
Tätä katsausta varten kävimme läpi huomattavan määrän kokeellisia tutkimuksia eri metsänkäsittely- ja luonnonhoitotoimien vaikutuksista monimuotoisuuteen. Tuloksia on parinkymmenen vuoden aikana saatu lukuisista eliöryhmistä, mutta emme voineet välttyä siltä vaikutelmalta, että tutkimukset ovat aiheiltaan kirjavia, lyhytaikaisella tutkimusrahoituksella toteutettuja, ja eri tutkimuskohteiden, kuten jokin tietty hakkuumenetelmä, osalta myös verraten vähäisiä. Havaitsimme seuraavia tiedollisia puutteita.
Metsikkötasolla tutkittavaa on edelleen paljon. Uutta, käytäntöön sovellettavissa olevaa tietoa tuottaisivat esimerkiksi vertailu ryhminä ja yksittäin (tasaisesti) jätettyjen säästöpuiden lajistovaikutuksista, tutkimus uudistusalan koon kynnysarvoista lajien säilymisen ja palautumisen kannalta sekä eri maanmuokkausmenetelmien lajistovaikutuksien vertailututkimukset. Lajiston suojelun näkökulmasta hyödyllisiä olisivat myös lehtojen, harjujen ja muiden harvinaisten metsäympäristöjen luonnonhoidon kokeelliset tarkastelut sekä tutkimus, joka kohdistuu lajien metsänhoitovasteita sääteleviin mekanismeihin ja luonnonhoitotoimien vaikutuksiin lajiston vuorovaikutussuhteisiin.
Maisematasolla tutkimusta voisi mielestämme suunnata nykyistä enemmän alueellisten metsärakenteiden ja luonnonhäiriöiden (esim. tuuli- tai patogeenituhot) merkitykseen lajiston häiriönsiedolle ja palautumiselle. Myös erirakenteismetsätalouden vaikutukset ekosysteemien toimintaan ja ekosysteemipalveluihin ovat heikosti tunnettuja, varsinkin alueellisella tasolla, vaikka tällaista metsätaloutta on viime aikoina julkisessa keskustelussa esitetty voimakkaasti lisättäväksi. Lopuksi on syytä nostaa esiin tarve pitkäaikaiselle seurantatutkimukselle ilmaston muuttuessa, esimerkkinä häiriöfrekvenssin (mm. metsätuhot) vaikutukset monimuotoisuuteen.
4.4 Ehdotuksia monimuotoisuuden huomioimiseksi metsätaloudessa
Tarkasteltaessa tuoreimpia aitoja uhanalaisuuden luokkamuutoksia havaitaan metsälajeilla lähes yhtä paljon positiivisia kuin negatiivisiakin muutoksia (109 ja 140; Hyvärinen ym. 2019). Tämä saattaa herättää toiveen, että uhanalaistumiskehitys metsissämme olisi pysähtynyt. Useat kymmenet positiivisen luokkamuutoksen lajit ovat kuitenkin eteläisten lehtimetsien perhosia ja kovakuoriaisia, jotka mitä ilmeisimmin hyötyvät ilmaston lämpenemisestä. Toisaalta esimerkiksi 51 metsäjäkälälajin – pääosin vanhoja metsiä tai kookkaita puita elinpaikoikseen vaativia – luokka oli heikentynyt ja vain kahden kohentunut. Ehdottamamme luonnonhoidon toimenpiteet (järeän kuolleen puun ja yksittäisten isokokoisten elävien puiden säästäminen sekä käsittelemämme luonnonhoitotoimet) todennäköisesti auttaisivat näiden lajien tilannetta metsiköistä maisematasolle. Talousmetsien luonnonhoidon laadulla on monimuotoisuuden säilymiselle ja lajiston palautumiselle ratkaiseva merkitys jo pelkästään siksi, että Etelä-Suomessa yli 90 % metsämaasta on talousmetsiä (Luonnonvarakeskus 2019).
Läpikäymiemme kokeellisten tutkimusten perusteella uhanalaisille lajeille näyttävät hakkuutapaa tärkeämpiä olevan tietyt metsän rakennepiirteet, kuten lahopuun määrä ja monipuolisuus sekä niiden jatkumoa turvaava isokokoinen säästöpuusto. Olemassa olevan lahopuuston säästäminen eri hakkuukäsittelyissä, mukaan lukien harvennukset ja maanmuokkaus, on kustannustehokkain tapa monimuotoisuuden ylläpitämiseksi. Myös metsikön pidennetty hakkuukierto näyttää tässä mielessä yhtä tehokkaalta kuin lahopuun keinollinen tuottaminen, koska elävät puut ja niiden myötä myös kuolevien puiden koko kasvavat (Ranius ym. 2005; Jonsson ym. 2006, 2010). Lisäksi säästöpuuston määrän kasvu metsikössä näkyy lahopuun määrän ja laadullisen vaihtelun kasvuna (Santaniello ym. 2016, 2017).
Korpipainanteiden aluskasvillisuus on lajistoltaan monimuotoisempaa kuin ympäröivissä kangasmetsissä (Vanha-Majamaa ja Jalonen 2001). Korpilaikuissa elää niihin erikoistunutta lajistoa, kuten epifyytti- ja epiksyylisammalia ja -jäkäliä sekä kovakuoriaisia, kuten uhanalainen korpikolva (Pytho kolwensis Sahlberg) (Siitonen ja Saaristo 2000). Näillä paikoilla on siten merkitystä lajiston säilymiselle uudistusvaiheen yli ja palautumiselle hakkuun jälkeen, joten ne tulisi jättää käsittelemättä varsinkin silloin, jos niissä on runsaasti lahopuuta. Tutkitun kaltaisia korpityyppejä turvaa Suomessa metsälaki, jonka listaamat avainbiotoopit tosin on kategorisesti määritelty pienialaisiksi ja/tai taloudellisesti vähäarvoisiksi (Suomen metsäkeskus 2018). Mönkkönen ym. (2011) osoittivat hehtaarin-parin avainbiotooppilaikkujen olevan monimuotoisuuden turvaamisen kannalta tehottomia. Tästä huolimatta pienemmiltäkin avainbiotoopeilta löytyy erikoistunutta lajistoa tavanomaista kangasmetsää enemmän (Wikberg ym. 2009), joten niiden säästäminen on lajiston säilymisen kannalta välttämätöntä (Häkkilä ym. 2020).
Kulotuksen käyttöä tulisi tutkimustulostemme perusteella lisätä alueilla, jotka luontaisestikin ilman tehokasta metsäpalojen torjuntaa palaisivat useasti, kuten harju- ja paahdeympäristöissä sekä mäntykankailla. Isojen metsäpaloalueiden suojelu voisi olla erittäin kustannustehokas tapa lisätä tärkeitä palonjälkeisiä eri sukkessiovaiheiden metsiä ja luoda palojatkumoa monimuotoisuuden ylläpitämiseksi.
Maisemarakenne voi vaikuttaa luonnonhoidon ja ennallistamisen onnistumiseen, joskin asia on huonosti tutkittu. Lajiston palautuminen kohteelle voi edellyttää lähialueiden lähdepopulaatioita ja niitä yhdistäviä levittäytymisreittejä, nk. ekologisia käytäviä (Hanski 2000; Hedenås ja Ericson 2008). Näin ollen runsaasti lähdealueita käsittävillä tai ympäristötyypeiltään ja metsärakenteiltaan rikkaammilla alueilla olisi paremmat monimuotoisuustoimien onnistumisen edellytykset. Joukko ruotsalaistutkimuksia osoitti, että lähiseudulla olevat suojelualueet eivät näkyneet tekopökkelöiden kovakuoriaislajistossa, mutta ympäröivien metsien puuston järeydellä ja puulajistolla oli siihen voimakas vaikutus (Lindbladh ja Abrahamsson 2008; Abrahamsson ym. 2009; Hallinger ym. 2018; Jonsell ym. 2019). Toisaalta reunametsän etäisyyden ei havaittu vaikuttavan sammallajien palautumiseen avohakkuun jälkeen (Hylander 2009).
Monimuotoisuutta hyödyttäisi myös sekapuustoisuusasteen nostaminen talousmetsissä. Tämä turvaa resursseja useammille lajeille kuin yhden puulajin metsiköt. Pieneltä osin ja lähes ilman kustannuksia tätä tavoitetta tukee taloudellisesti vähäarvoisten puiden säästäminen hakkuissa. Sekapuustrategia on myös metsäteollisuuden tulevaisuuden toimintaedellytyksien kannalta intuitiivisesti järkevä muuttuvassa ilmastossa.
Kiitokset
Toimitus ja kaksi ulkopuolista arvioijaa esittivät joukon hyödyllisiä kommentteja, joiden huomioiminen paransi käsikirjoitusta merkittävästi. Parhaat kiitoksemme.
Lähteet
Abrahamsson M, Lindbladh M, Rönnberg J (2008) Influence of butt rot on beetle diversity in artificially created high-stumps of Norway spruce. For Ecol Manage 255: 3396–3403. https://doi.org/10.1016/j.foreco.2008.01.010.
Abrahamsson M, Jonsell M, Niklasson M, Lindbladh M (2009) Saproxylic beetle assemblages in artificially created high-stumps of spruce (Picea abies) and birch (Betula pendula/pubescens) – does the surrounding landscape matter? Insect Conserv Divers 2: 284–294. https://doi.org/10.1111/j.1752-4598.2009.00066.x.
Andersson J, Hjältén J, Dynesius M (2015) Wood-inhabiting beetles in low stumps, high stumps and logs on boreal clear-cuts: implications for dead wood management. PLoS One 10, article id e0118896. https://doi.org/10.1371/journal.pone.0118896.
Bader P, Jansson S, Jonsson BG (1995) Wood-inhabiting fungi and substratum decline in selectively logged boreal spruce forests. Biol Conserv 72: 355–362. https://doi.org/10.1016/0006-3207(94)00029-P.
Beese WJ, Deal J, Dunsworth BG, Mitchell SJ, Philpot T (2019) Two decades of variable retention in British Columbia: a review of its implementation and effectiveness for biodiversity conservation. Ecol Process 8, article id 33. https://doi.org/10.1186/s13717-019-0181-9.
Berglund H, Jönsson MT, Penttilä R, Vanha-Majamaa I (2011) The effects of burning and dead-wood creation on the diversity of pioneer wood-inhabiting fungi in managed boreal spruce forests. For Ecol Manage: 1293–1305. https://doi.org/10.1016/j.foreco.2011.01.008.
Djupström L, Weslien J (2019) Effaråsen – att bruka och bevara i gammal tallskog. SkogForsk Arbetsrapport 1009-2019.
Drössler L (2016) Heterogen skog – för komplex för skogsbruk och tillämpad skogsforskning? Rön för Sveriges Lantbruksuniversitet Nr 9.
Edenius L, Elmberg J (1996). Landscape level effects of modern forestry on bird communities in North Swedish boreal forests. Landscape Ecol 11: 325–338. https://doi.org/10.1007/BF02447520.
Elo M, Halme P, Toivanen T, Kotiaho JS (2019) Species richness of polypores can be increased by supplementing dead wood resource into a boreal forest landscape. J Appl Ecol 56: 1267–1277. https://doi.org/10.1111/1365-2664.13364.
Eriksson AM, Olsson J, Jonsson BG, Toivanen S, Edman M (2013) Effects of restoration fire on dead wood heterogeneity and availability in three Pinus sylvestris forests in Sweden. Silva Fenn 47, article id 954. https://doi.org/10.14214/sf.954.
Eriksson M, Lilja S, Roininen H (2006) Dead wood creation and restoration burning: implications for bark beetles and beetle induced tree deaths. For Ecol Manage 231: 205–213. https://doi.org/10.1016/j.foreco.2006.05.050.
Euroopan Unioni (2020). EU biodiversity strategy for 2030. https://ec.europa.eu/environment/nature/biodiversity/strategy/index_en.htm. Viitattu 10.11.2020.
Fedrowitz K, Koricheva J, Baker SC, Lindenmayer DB, Palik B, Rosenvald R, Beese W, Franklin JF, Kouki J, Macdonald E, Messier C, Sverdrup-Thygeson A, Gustafsson L (2014) Can retention forestry help conserve biodiversity? A meta-analysis. J Appl Ecol 51: 1669–1679. https://doi.org/10.1111/1365-2664.12289.
Fossestol KO, Sverdrup-Thygeson A (2009) Saproxylic beetles in high stumps and residual downed wood on clear-cuts and in forest edges. Scand J For Res 24: 403–416. https://doi.org/10.1080/02827580903143871.
Gibb H, Pettersson RB, Hjältén J, Hilszczanski J, Ball JP, Johansson T, Atlegrim O, Danell K (2006) Conservation-oriented forestry and early successional saproxylic beetles: responses of functional groups to manipulated dead wood substrates. Biol Conserv 129: 437–450. https://doi.org/10.1016/j.biocon.2005.11.010.
Granath G, Kouki J, Johnson S, Heikkala O, Rodríguez A, Strengbom J (2018) Trade-offs in berry production and biodiversity under prescribed burning and retention regimes in boreal forests. J Appl Ecol 55: 1658–1667. https://doi.org/10.1111/1365-2664.13098.
Gustafsson L, Berglind M, Granström A, Grelle A, Isacsson G, Kjellander P, Larsson S, Lindh M, Pettersson L.B, Strengbom J, Stridh B, Sävström T, Thor G, Wikars L-O, Mikusiński G (2019) Rapid ecological response and intensified knowledge accumulation following a north European mega-fire. Scand J For Res 34: 234–253. https://doi.org/10.1080/02827581.2019.1603323.
Götmark F, Paltto H, Nordén B, Götmark E (2005) Evaluating partial cutting in broadleaved temperate forest under strong experimental control: short-term effects on herbaceous plants. For Ecol Manage 214: 124–141. https://doi.org/10.1016/j.foreco.2005.03.05.
Hagner M (1992) Biologiskt och ekonomiskt resultat i fältförsök med plockhuggning kombinerad med plantering. Arbetsrapport 63, Sveriges Lantbruksuniversitet, Umeå.
Hallinger M, Kärvemo S, Ranius T (2018) Does it pay to concentrate conservation efforts for dead-wood dependent insects close to existing reserves: a test on conservation planning in Sweden. Insect Conservation and Diversity 11: 317–329. https://doi.org/10.1111/icad.12279.
Hanski I (2000) Extinction debt and species credit in boreal forests: modelling the consequences of different approaches to biodiversity conservation. Annales Zoologici Fennici 37: 271–280. https://www.jstor.org/stable/23735720.
Hautala H, Vanha-Majamaa I (2006) Immediate tree uprooting after retention-felling in a coniferous boreal forest in Fennoscandia. Can J For Res 36: 3167–3172. https://doi.org/10.1139/x06-193.
Hautala H, Jalonen J, Laaka-Lindberg S, Vanha-Majamaa I (2004) Impacts of retention felling on coarse woody debris (CWD) in mature boreal spruce forests in Finland. Biodivers Conserv 13: 1541–1554. https://doi.org/10.1023/B:BIOC.0000021327.43783.a9.
Hautala H, Laaka-Lindberg S, Vanha-Majamaa I (2011) Effects of retention felling on epixylic species in boreal spruce forests in southern Finland. Restor Ecol 19: 418–429. https://doi.org/10.1111/j.1526-100X.2009.00545.x.
Hedenås H, Ericson L (2003) Response of epiphytic lichens on Populus tremula in a selective cutting experiment. Ecol Appl 13: 1124–1134. https://doi.org/10.1890/1051-0761(2003)13[1124:ROELOP]2.0.CO;2.
Hedenås H, Ericson L (2008) Species occurrences at stand level cannot be understood without considering the landscape context: cyanolichens on aspen in boreal Sweden. Biol Conserv 141: 710–718. https://doi.org/10.1016/j.biocon.2007.12.019.
Hedwall P-O, Brunet J, Nordin A, Bergh J (2013) Changes in the abundance of keystone forest floor species in response to changes of forest structure. J Veg Sci 24: 296–306. https://doi.org/10.1111/j.1654-1103.2012.01457.x.
Heikkala O, Suominen M, Junninen K, Hämäläinen A, Kouki J (2014) Effects of retention level and fire on retention tree dynamics in boreal forests. For Ecol Manage 328: 193–201. https://doi.org/10.1016/j.foreco.2014.05.022.
Heikkala O, Martikainen P, Kouki J (2016a) Decadal effects of emulating natural disturbances in forest management on saproxylic beetle assemblages. Biol Conserv 194: 39–47. https://doi.org/10.1016/j.biocon.2015.12.002.
Heikkala O, Seibold S, Koivula M, Martikainen P, Thorn S, Müller J, Kouki J (2016b) Retention forestry and prescribed burning result in functionally different saproxylic beetle assemblages than clear-cutting. For Ecol Manage 359: 51–58. https://doi.org/10.1016/j.foreco.2015.09.043.
Heikkala O, Martikainen P, Kouki J (2017) Prescribed burning is an effective and quick method to conserve rare pyrophilous forest-dwelling flat bugs. Insect Conserv Diver 10: 32–41. https://doi.org/10.1111/icad.12195.
Hekkala A-M, Päätalo M-L, Tarvainen O, Tolvanen A (2014a) Restoration of young forests in eastern Finland: benefits for saproxylic beetles (Coleoptera). Restor Ecol 22: 151–159. https://doi.org/10.1111/rec.12050.
Hekkala A-M, Tarvainen O, Tolvanen A (2014b) Dynamics of understory vegetation after restoration of natural characteristics in the boreal forests in Finland. For Ecol Manage 330: 55–66. https://doi.org/10.1016/j.foreco.2014.07.001.
Hekkala A-M, Ahtikoski A, Päätalo M-L, Tarvainen O, Siipilehto J, Tolvanen A (2016) Restoring volume, diversity and continuity of deadwood in boreal forests. Biodivers Conserv 25: 1107–1132. https://doi.org/10.1007/s10531-016-1112-z.
Hjältén J, Stenbacka F, Andersson J (2010) Saproxylic beetle assemblages on low stumps, high stumps and logs: implications for environmental effects of stump harvesting. For Ecol Manage 260: 1149–1155. https://doi.org/10.1016/j.foreco.2010.07.003.
Hyvärinen E, Kouki J, Martikainen P, Lappalainen H (2005) Short-term effects of controlled burning and green-tree retention on beetle (Coleoptera) assemblages in managed boreal forests. For Ecol Manage 212: 315–332. https://doi.org/10.1016/j.foreco.2005.03.029.
Hyvärinen E, Kouki J, Martikainen P (2006) Fire and green-tree retention in conservation of red-listed and rare deadwood-dependent beetles in Finnish boreal forests. Conserv Biol 20: 1711–1719. https://doi.org/10.1111/j.1523-1739.2006.00511.x.
Hyvärinen E, Kouki J, Martikainen P (2009) Prescribed fires and retention trees help to conserve beetle diversity in managed boreal forests despite their transient negative effects on some beetle groups. Insect Conserv Diver 2: 93–105. https://doi.org/10.1111/j.1752-4598.2009.00048.x.
Hyvärinen E, Juslén A, Kemppainen E, Uddström A, Liukko U-M (toim) (2019) Suomen lajien uhanalaisuus 2019. Ympäristöministeriö ja Suomen Ympäristökeskus, Helsinki. http://hdl.handle.net/10138/299501. Viitattu 24.2.2021.
Hägglund R, Hjältén J (2018) Substrate specific restoration promotes saproxylic beetle diversity in boreal forest set-asides. For Ecol Manage 425: 45–58. https://doi.org/10.1016/j.foreco.2018.05.019.
Hägglund R, Hekkala A-M, Hjältén J, Tolvanen A (2015) Positive effects of ecological restoration on rare and threatened flat bugs (Heteroptera: Aradidae). J Insect Conserv 19: 1089–1099. https://doi.org/10.1007/s10841-015-9824-z.
Häkkilä M, Savilaakso S, Johansson A, Sandgren T, Uusitalo A, Mönkkönen M, Puttonen P (2020) Pienialaisten suojeltujen elinympäristöjen merkitys luonnon monimuotoisuudelle boreaalisissa talousmetsissä. Systemaattinen kirjallisuuskatsaus. Metsätehon raportti 257. https://www.metsateho.fi/wp-content/uploads/Raportti_257_Pienialaisten_suojeltujen_elinymparistojen.pdf. Viitattu 24.2.2021.
Hämäläinen A, Kouki J, Lõhmus P (2014) The value of retained Scots pines and their dead wood legacies for lichen diversity in clear-cut forests: the effects of retention level and prescribed burning. For Ecol Manage 324: 89–100. https://doi.org/10.1016/j.foreco.2014.04.016.
Hämäläinen A, Hujo M, Heikkala O, Junninen K, Kouki J (2016) Retention tree characteristics have major influence on the post-harvest tree mortality and availability of coarse woody debris in clear-cut areas. For Ecol Manage 369: 66–73. https://doi.org/10.1016/j.foreco.2016.03.037.
Joelsson K, Hjältén J, Work T, Gibb H, Roberge J-M, Löfroth T (2017) Uneven-aged silviculture can reduce negative effects of forest management on beetles. For Ecol Manage 391: 436–445. https://doi.org/10.1016/j.foreco.2017.02.006.
Joelsson K, Hjältén J, Gibb H (2018) Forest management strategy affects saproxylic beetle assemblages: a comparison of even and uneven-aged silviculture using direct and indirect sampling. PLoS One 13, article id e0194905. https://doi.org/10.1371/journal.pone.0194905.
Johansson T, Gibb H, Hilszczanski J, Pettersson RB, Hjältén J, Atlegrim O, Ball JP, Danell K (2006) Conservation-oriented manipulations of coarse woody debris affect its value as habitat for spruce-infesting bark and ambrosia beetles (Coleoptera: Scolytinae) in northern Sweden. Can J For Res 36: 174–185. https://doi.org/10.1139/x05-235.
Johansson T, Hjältén J, Stenbacka F, Dynesius M (2010) Responses of eight boreal flat bug (Heteroptera: Aradidae) species to clear-cutting and forest fire. J Insect Conserv 14: 3–9. https://doi.org/10.1007/s10841-009-9218-1.
Johnson S, Strengbom J, Kouki J (2014) Low levels of tree retention do not mitigate the effects of clearcutting on ground vegetation dynamics. For Ecol Manage 330: 67–74. https://doi.org/10.1016/j.foreco.2014.06.031.
Jokela J, Siitonen J, Koivula M (2019) Short-term effects of selection, gap, patch and clear cutting on the beetle fauna in boreal spruce-dominated forests. For Ecol Manage 446: 29–37. https://doi.org/10.1016/j.foreco.2019.05.027.
Jonsell M, Hansson J (2011) Logs and stumps in clearcuts support similar saproxylic beetle diversity: implications for bioenergy harvest. Silva Fenn 45: 1053–1064. https://doi.org/10.14214/sf.86.
Jonsell M, Nittérus K, Stighäll K (2004) Saproxylic beetles in natural and man-made deciduous high stumps retained for conservation. Biol Conserv 118: 163–173. https://doi.org/10.1016/j.biocon.2003.08.017.
Jonsell M, Schroeder M, Weslien J (2005) Saproxylic beetles in high stumps of spruce: fungal flora important for determining the species composition. Scand J For Res 20: 54–62. https://doi.org/10.1080/02827580510008211.
Jonsell M, Abrahamsson M, Widenfalk L (2019) Increasing influence of the surrounding landscape on saproxylic beetle communities over 10 years succession in dead wood. For Ecol Manage 440: 267–284. https://doi.org/10.1016/j.foreco.2019.02.021.
Jonsson M, Ranius T, Ekvall H, Bostedt G, Dahlberg A, Ehnström B, Nordén B, Stokland JN (2006) Cost-effectiveness of silvicultural measures to increase substrate availability for red-listed wood-living organisms in Norway spruce forests. Biol Conserv 127: 443–462. https://doi.org/10.1016/j.biocon.2005.09.004.
Jonsson M, Ranius T, Ekvall H, Bostedt G (2010) Cost-effectiveness of silvicultural measures to increase substrate availability for wood-dwelling species: a comparison among boreal tree species. Scand J For Res 25: 46–60. https://doi.org/10.1080/02827581003620347.
Junninen K, Komonen A (2011) Conservation ecology of boreal polypores: a review. Biol Conserv 144: 11–20. https://doi.org/10.1016/j.biocon.2010.07.010.
Junninen K, Penttilä R, Martikainen P (2007) Fallen retention aspen trees on clear-cuts can be important habitats for red-listed polypores: a case study in Finland. Biodivers Conserv 16: 475–490. https://doi.org/10.1007/s10531-005-6227-6.
Junninen K, Kouki J, Renvall P (2008) Restoration of natural legacies of fire in European boreal forests: an experimental approach to the effects on wood-decaying fungi. Can J For Res 38: 202–215. https://doi.org/10.1139/X07-145.
Kaila L, Martikainen P, Punttila P (1997) Dead trees left in clear-cuts benefit saproxylic Coleoptera adapted to natural disturbances in boreal forest. Biodivers Conserv 6: 1–18. https://doi.org/10.1023/A:1018399401248.
Kaila S (toim) (1998) Monimuotoisuus talousmetsän uudistamisessa -hankkeen väliraportit (MONTA-hanke). Metsätehon raportti 62. http://www.metsateho.fi/wp-content/uploads/2015/02/metsatehon_raportti_062.pdf. Viitattu 10.11.2020.
Koivula M (2002) Alternative harvesting methods and boreal carabid beetles (Coleoptera, Carabidae). For Ecol Manage 167: 103–121. https://doi.org/10.1016/S0378-1127(01)00717-4.
Koivula M, Vanha-Majamaa I (2020) Experimental evidence on biodiversity impacts of variable retention forestry, prescribed burning, and deadwood manipulation in Fennoscandia. Ecol Process 9, article id 11. https://doi.org/10.1186/s13717-019-0209-1.
Koivula M, Kuuluvainen T, Hallman E, Kouki J, Siitonen J, Valkonen S (2014) Forest management inspired by natural disturbance dynamics (DISTDYN) - a long-term research and development project in Finland. Scand J For Res 29: 579–592. https://doi.org/10.1080/02827581.2014.938110.
Koivula MJ, Venn S, Hakola P, Niemelä J (2019) Responses of boreal ground beetles (Coleoptera, Carabidae) to different logging regimes ten years post-harvest. For Ecol Manage 436: 27–38. https://doi.org/10.1016/j.foreco.2018.12.047.
Komonen A, Halme P, Jäntti M, Koskela T, Kotiaho J, Toivanen T (2014) Created substrates do not fully mimic natural substrates in restoration: the occurrence of polypores on spruce logs. Silva Fenn 48, article id 980. https://doi.org/10.14214/sf.980.
Kontula T, Raunio A (toim) (2018) Suomen luontotyyppien uhanalaisuus 2018. Luontotyyppien punainen kirja. Suomen Ympäristö 5/2018.
Korhonen KT, Ihalainen A, Ahola A, Heikkinen J, Henttonen HM, Hotanen J-P, Nevalainen S, Pitkänen J, Strandström M, Viiri H (2017) Suomen metsät 2009–2013 ja niiden kehitys 1921–2013. Luonnonvara- ja biotalouden tutkimus 59/2017, Luonnonvarakeskus, Helsinki. http://urn.fi/URN:ISBN:978-952-326-467-0.
Kotiaho JS, Kuusela S, Nieminen E, Päivinen J (toim) (2015) Elinympäristöjen tilan edistäminen Suomessa. ELITE-työryhmän mietintö elinympäristöjen tilan edistämisen priorisointisuunnitelmaksi ja arvio suunnitelman kokonaiskustannuksista. Suomen Ympäristö 8/2015. http://hdl.handle.net/10138/156982.
Kruys N, Fridman J, Götmark F, Simonsson P, Gustafsson L (2013) Retaining trees for conservation at clearcutting has increased structural diversity in young Swedish production forests. For Ecol Manage 304: 312–321. https://doi.org/10.1016/j.foreco.2013.05.018.
Kuuluvainen T, Grenfell R (2012) Natural disturbance emulation in boreal forest ecosystem management – theories, strategies, and a comparison with conventional even-aged management. Can J For Res 42: 1185–1203. https://doi.org/10.1139/x2012-064.
Kuuluvainen T, Lindberg H, Vanha-Majamaa I, Keto-Tokoi P, Punttila P (2019) Low-level retention forestry, certification and biodiversity: case Finland. Ecol Process 8: 47. https://doi.org/10.1186/s13717-019-0198-0.
Kärvemo S, Björkman C, Johansson T, Weslien J, Hjältén J (2017) Forest restoration as a double-edged sword: the conflict between biodiversity conservation and pest control. J Appl Ecol 54: 1658–1668. https://doi.org/10.1111/1365-2664.12905.
Larsson S (2018) Forest fires in Sweden 2014 and 2018. Forest industry guidelines to prevent forest fires. Barents Forest Sector Network, Skogsstyrelsen.
Lehtonen I, Venäläinen A (2020) Metsäpalokesä 2018 muuttuvassa ilmastossa – poikkeuksellinen vuosi vai uusi normaali? Raportteja 2020:2, Ilmatieteen laitos. http://hdl.handle.net/10138/315097.
Lilja S, De Chantal M, Kuuluvainen T, Vanha-Majamaa I, Puttonen P (2005) Restoring natural characteristics in managed Norway spruce [Picea abies (L.) Karst.] stands with partial cutting, dead wood creation and fire: immediate treatment effects. Scand J For Res 20: 68–78. https://doi.org/10.1080/14004080510040977.
Lindberg H, Punttila P, Vanha-Majamaa I (2020) The challenge of combining variable retention and prescribed burning in Finland. Ecol Process 9, article id 1. https://doi.org/10.1186/s13717-019-0207-3.
Lindbladh M, Abrahamsson M (2008) Beetle diversity in high-stumps from Norway spruce thinnings. Scand J For Res 23: 339–347. https://doi.org/10.1080/02827580802282762.
Lindbladh M, Abrahamsson M, Seedre M, Jonsell M (2007) Saproxylic beetles in artificially created high-stumps of spruce and birch within and outside hotspot areas. Biodivers Conserv 16: 3213–3226. https://doi.org/10.1007/s10531-007-9173-7.
Lindhe A, Lindelöw Å (2004) Cut high stumps of spruce, birch, aspen and oak as breeding substrates for saproxylic beetles. For Ecol Manage 203: 1–20. https://doi.org/10.1016/j.foreco.2004.07.047.
Lindhe A, Åsenblad N, Toresson H-G (2004) Cut logs and high stumps of spruce, birch, aspen and oak – nine years of saproxylic fungi succession. Biol Conserv 119: 443–454. https://doi.org/10.1016/j.biocon.2004.01.005.
Lindhe A, Lindelöw Å, Åsenblad N (2005) Saproxylic beetles in standing dead wood density in relation to substrate sun-exposure and diameter. Biodivers Conserv 14: 3033–3053. https://doi.org/10.1007/s10531-004-0314-y.
Lõhmus P, Leppik E, Motiejunaite J, Suija A, Lõhmus A (2012) Old selectively cut forests can host rich lichen communities – lessons from an exhaustive field survey. Nova Hedwigia 95: 493–515. https://doi.org/10.1127/0029-5035/2012/0064.
Luonnonvarakeskus (2016) Metsävarat. https://www.luke.fi/tietoa-luonnonvaroista/metsa/metsavarat-ja-metsasuunnittelu/metsavarat/. Viitattu 10.11.2020.
Luonnonvarakeskus (2019) Metsien suojelu 1.1.2019. https://stat.luke.fi/metsien-suojelu-112019_fi. Viitattu 10.11.2020.
Martikainen P (2001) Conservation of threatened saproxylic beetles: significance of retained aspen Populus tremula on clearcut areas. Ecol Bull 49: 205–218.
Martikainen P, Kouki J, Heikkala O (2006a) The effects of green tree retention and subsequent prescribed burning on ground beetles (Coleoptera: Carabidae) in boreal pine-dominated forests. Ecography 29: 659–670. https://doi.org/10.1111/j.2006.0906-7590.04562.x.
Martikainen P, Kouki J, Heikkala O, Hyvärinen E, Lappalainen H (2006b) Effects of green tree retention and prescribed burning on the crown damage caused by the pine shoot beetles (Tomicus spp.) in pine-dominated timber harvest areas. J Appl Entomol 130: 37–44. https://doi.org/10.1111/j.1439-0418.2005.01015.x.
Matveinen-Huju K, Koivula M (2008) Effects of alternative harvesting methods on boreal forest spider assemblages. Can J For Res 38: 782–794. https://doi.org/10.1139/X07-169.
Matveinen-Huju K, Niemelä J, Rita H, O’Hara RB (2006) Retention-tree groups in clear-cuts: do they constitute ‘life-boats’ for spiders and carabids? For Ecol Manage 230: 119–135. https://doi.org/10.1016/j.foreco.2006.04.025.
Mönkkönen M, Reunanen P, Kotiaho JS, Juutinen A, Tikkanen O-P, Kouki J (2011) Cost-effective strategies to conserve boreal forest biodiversity and long-term landscape-level maintenance of habitats. Eur J For Res 130: 717–727. https://doi.org/10.1007/s10342-010-0461-5.
Norberg A, Halme P, Kotiaho JS, Toivanen T, Ovaskainen O (2019) Experimentally induced community assembly of polypores reveals the importance of both environmental filtering and assembly history. Fungal Ecol 41: 137–146. https://doi.org/10.1016/j.funeco.2019.05.003.
Nordén B, Götmark F, Ryberg M, Paltto H, Allmér J (2008) Partial cutting reduces species richness of fungi on woody debris in oak-rich forests. Can J For Res 38: 1807–1816. https://doi.org/10.1139/X08-031.
Nordén B, Paltto H, Claesson C, Götmark F (2012) Partial cutting can enhance epiphyte conservation in temperate oak-rich forests. For Ecol Manage 270: 35–44. https://doi.org/10.1016/j.foreco.2012.01.014.
Oldén A, Selonen VAO, Lehkonen E, Kotiaho JS (2019a) The effect of buffer strip width and selective logging on streamside plant communities. BMC Ecol 19, article id 9. https://doi.org/10.1186/s12898-019-0225-0.
Oldén A, Peura M, Saine S, Kotiaho JS, Halme P (2019b) The effect of buffer strip width and selective logging on riparian forest microclimate. For Ecol Manage 453, article id 117623. https://doi.org/10.1016/j.foreco.2019.117623.
Olsson J, Jonsson BG, Hjältén J, Ericson L (2011) Addition of coarse woody debris – the early fungal succession on Picea abies logs in managed forests. Biol Conserv 144: 1100–1110. https://doi.org/10.1016/j.biocon.2010.12.029.
Paltto H, Nordén B, Götmark F (2008) Partial cutting as a conservation alternative for oak (Quercus spp.) forest – responses of bryophytes and lichens on dead wood. For Ecol Manage 256: 536–547. https://doi.org/10.1016/j.foreco.2008.04.045.
Pasanen H (2017) Ecological effects of disturbance-based restoration in boreal forests. Dissertationes Forestales 244. https://doi.org/10.14214/df.244.
Pasanen H, Junninen K, Kouki J (2014) Restoring dead wood in forests diversifies wood-decaying fungal assemblages but does not quickly benefit red-listed species. For Ecol Manage 312: 92–100. https://doi.org/10.1016/j.foreco.2013.10.018.
Pasanen H, Junninen K, Boberg J, Tatsumi S, Stenlid J, Kouki J (2018) Life after tree death: does restored dead wood host different fungal communities to natural woody substrates? For Ecol Manage 409: 863–871. https://doi.org/10.1016/j.foreco.2017.12.021.
Pasanen H, Juutilainen K, Siitonen J (2019) Responses of polypore fungi following disturbance-emulating harvesting treatments and deadwood creation in boreal Norway spruce dominated forests. Scand J For Res 34: 557–568. https://doi.org/10.1080/02827581.2019.1663915.
Perhans K, Appelgren L, Jonsson F, Nordin U, Söderström B, Gustafsson L (2009) Retention patches as potential refugia for bryophytes and lichens in managed forest landscapes. Biol Conserv 142: 1125–1133. https://doi.org/10.1016/j.biocon.2008.12.033.
Pietikäinen AS, Haimi J, Siitonen J (2003) Short-term responses of soil macroarthropod community to clear felling and alternative forest regeneration methods. For Ecol Manage 172: 339–353. https://doi.org/10.1016/S0378-1127(01)00811-8.
Pihlaja M, Koivula M, Niemelä J (2006) Responses of boreal carabid beetle assemblages (Coleoptera, Carabidae) to clear-cutting and top-soil preparation. For Ecol Manage 222: 182–190. https://doi.org/10.1016/j.foreco.2005.10.013.
Rabinowitsch-Jokinen R, Vanha-Majamaa I (2010) Immediate effects of logging, mounding and removal of logging residues and stumps on coarse woody debris in managed boreal Norway spruce stands. Silva Fenn 44: 51–61. https://doi.org/10.14214/sf.162.
Rabinowitsch-Jokinen R, Laaka-Lindberg S, Vanha-Majamaa I (2012) Immediate effects of logging, mounding and removal of logging residues on epixylic species in managed boreal Norway spruce stands in Southern Finland. J Sustain Forest 31: 205–229. https://doi.org/10.1080/10549811.2011.582825.
Ramberg E, Strengbom J, Granath G (2018) Coordination through databases can improve prescribed burning as a conservation tool to promote forest biodiversity. Ambio 47: 298–306. https://doi.org/10.1007/s13280-017-0987-6.
Ranius T, Ekvall H, Jonsson M, Bostedt G (2005) Cost-efficiency of measures to increase the amount of coarse woody debris in managed Norway spruce forests. For Ecol Manage 206: 119–133. https://doi.org/10.1016/j.foreco.2004.10.061.
Rodriguez A, Kouki J (2017) Disturbance-mediated heterogeneity drives pollinator diversity in boreal managed forest ecosystems. Ecol Appl 27: 589–602. https://doi.org/10.1002/eap.1468.
Rudolphi J, Caruso A, von Cräutlein M, Laaka-Lindberg S, Ryömä R, Berglund H (2011) Relative importance of thinned and clear-cut stands for bryophyte diversity on stumps. For Ecol Manage 261: 1911–1918. https://doi.org/10.1016/j.foreco.2011.02.014.
Saaristo L, Vanhatalo K (toim) (2019) Metsänhoidon suositukset talousmetsien luonnonhoitoon, työopas. Tapion julkaisuja.
Sandström J, Bernes C, Junninen K, Lõhmus A, Macdonald E, Müller J, Jonsson BG (2019) Impacts of dead wood manipulation on the biodiversity of temperate and boreal forests: a systematic review. J Appl Ecol 56: 1770–1781. https://doi.org/10.1111/1365-2664.13395.
Santaniello F, Djupström LB, Ranius T, Rudolphi J, Widenfalk O, Weslien J (2016) Effects of partial cutting on logging productivity, economic returns and dead wood in boreal pine forest. For Ecol Manage 365: 152–158. https://doi.org/10.1016/j.foreco.2016.01.033.
Santaniello F, Djupström LB, Ranius T, Weslien J, Rudolphi J, Thor G (2017) Large proportion of wood dependent lichens in boreal pine forest are confined to old hard wood. Biodivers Conserv 26: 1295–1310. https://doi.org/10.1007/s10531-017-1301-4.
Seidl R, Thom D, Kautz M, Martin-Benito D, Peltoniemi M, Vacchiano G, Wild J, Ascoli D, Petr M, Honkaniemi J, Lexer MJ, Trotsiuk V, Mairota P, Svoboda M, Fabrika M, Nagel TA, Reyer CPO (2017) Forest disturbances under climate change. Nat Clim Chang 7: 395–402. https://doi.org/10.1038/nclimate3303.
Selonen VAO, Kotiaho J (2013) Buffer strips can pre-empt extinction debt in boreal streamside habitats. BMC Biol 13, article id 24. https://doi.org/10.1186/1472-6785-13-24.
Siira-Pietikäinen A, Haimi J (2009) Changes in soil fauna 10 years after forest harvestings: comparison between clear felling and green-tree retention methods. For Ecol Manage 258: 332–338. https://doi.org/10.1016/j.foreco.2009.04.024.
Siitonen J, Saaristo L (2000) Habitat requirements and conservation of Pytho kolwensis, a beetle species of old-growth boreal forest. Biol Conserv 94: 211–220. https://doi.org/10.1016/S0006-3207(99)00174-3.
Sippola A-L, Similä M, Mönkkönen M, Jokimäki J (2004) Diversity of polyporous fungi (Polyporaceae) in northern boreal forests: effects of forest site type and logging intensity. Scand J For Res 19: 152–163. https://doi.org/10.1080/02827580410026294.
Storaunet KO, Rolstad J, Toeneiet M, Rolstad E (2008) Effect of logging on the threatened epiphytic lichen Usnea longissima: a comparative and retrospective approach. Silva Fenn 42: 685–703. https://doi.org/10.14214/sf.465.
Storaunet KO, Rolstad J, Rolstad E (2014) Effects of logging on the threatened epiphytic lichen Usnea longissima: an experimental approach. Silva Fenn 48, article id 949. https://doi.org/10.14214/sf.949.
Suomen metsäkeskus (2018) Tulkintasuosituksia metsälain 10§:n tarkoittamien erityisen tärkeiden elinympäristöjen rajaamisesta ja käsittelystä. https://www.metsakeskus.fi/sites/default/files/metsalain-10-pykala-kohteiden-tulkintasuositus.pdf. Viitattu 11.2.2021.
Suominen M, Junninen K, Heikkala O, Kouki J (2015) Combined effects of retention forestry and prescribed burning on polypore fungi. J Appl Ecol 52: 1001–1008. https://doi.org/10.1111/1365-2664.12447.
Suominen M, Junninen K, Heikkala O, Kouki J (2018) Burning harvested sites enhances polypore diversity on stumps and slash. For Ecol Manage 414: 47–53. https://doi.org/10.1016/j.foreco.2018.02.007.
Suominen M, Junninen K, Kouki J (2019) Diversity of fungi in harvested forests 10 years after logging and burning: polypore assemblages on different woody substrates. For Ecol Manage 446: 63–70. https://doi.org/10.1016/j.foreco.2019.05.030.
Sverdrup-Thygeson A, Ims RA (2002) The effect of forest clearcutting in Norway on the community of saproxylic beetles on aspen. Biol Conserv 106: 347–357. https://doi.org/10.1016/S0006-3207(01)00261-0.
Tapio Oy (2019) Metsänhoidon suositukset. https://www.metsanhoitosuositukset.fi/. Viitattu 10.11.2020.
Toivanen T, Kotiaho J (2007a) Mimicking natural disturbances of boreal forests: the effects of controlled burning and creating dead wood on beetle diversity. Biodivers Conserv 16: 3193–3211. https://doi.org/10.1007/s10531-007-9172-8.
Toivanen T, Kotiaho JS (2007b) Burning of logged sites to protect beetles in managed boreal forests. Conserv Biol 21: 1562–1572. https://doi.org/10.1111/j.1523-1739.2007.00808.x.
Toivanen T, Kotiaho J (2010) The preferences of saproxylic beetle species for different dead wood types created in forest restoration treatments. Can J For Res 40: 445–464. https://doi.org/10.1139/X09-205.
Toivanen T, Selonen V, Kotiaho JS (2004) Effects of management and restoration on forest biodiversity: an experimental approach. EFI Proceedings 51: 493–503. http://urn.fi/URN:NBN:fi:jyu-201905022384.
Toivanen T, Liikanen V, Kotiaho JS (2009). Effects of forest restoration treatments on the abundance of bark beetles in Norway spruce forests of southern Finland. For Ecol Manage 257: 117–125. https://doi.org/10.1016/j.foreco.2008.08.025.
Toivanen T, Heikkilä T, Koivula MJ (2014) Emulating natural disturbances in boreal Norway spruce forests: effects on ground beetles (Coleoptera, Carabidae). For Ecol Manage 314: 64–74. https://doi.org/10.1016/j.foreco.2013.11.028.
Tonteri T, Salemaa M, Rautio P, Hallikainen V, Korpela L, Merilä P (2016) Forest management regulates temporal change in the cover of boreal plant species. For Ecol Manage 381: 115–124. https://doi.org/10.1016/j.foreco.2016.09.015.
Tullus T, Rosenvald R, Leis M, Lõhmus P (2018) Impacts of shelterwood logging on forest bryoflora: distinct assemblages with richness comparable to mature forests. For Ecol Manage 411: 67–74. https://doi.org/10.1016/j.foreco.2018.01.008.
Tullus T, Tishler M, Rosenvald R, Tullus A, Lutter R, Tullus H (2019) Early responses of vascular plant and bryophyte communities to uniform shelterwood cutting in hemiboreal Scots pine forests. For Ecol Manage 440: 70–78. https://doi.org/10.1016/j.foreco.2019.03.009.
Vanha-Majamaa I, Jalonen J (2001) Green tree retention in Fennoscandian forestry. Scand J For Res Suppl. 3: 79–90. https://doi.org/10.1080/028275801300004433.
Vanha-Majamaa I, Reinikainen A (2000) Muuttuvan maankäytön vaikutus kasvillisuuteen. Julkaisussa: Reinikainen A, Mäkipää R, Vanha-Majamaa I, Hotanen JP (toim) Kasvit muuttuvassa metsäluonnossa. Tammi, Jyväskylä, ss 302–317.
Vanha-Majamaa I, Lilja S, Ryömä R, Kotiaho JS, Laaka-Lindberg S, Lindberg H, Puttonen P, Tamminen P, Toivanen T, Kuuluvainen T (2007) Rehabilitating boreal forest structure and species composition in Finland through logging, dead wood creation and fire: the EVO experiment. For Ecol Manage 250: 77–88. https://doi.org/10.1016/j.foreco.2007.03.012.
Vanha-Majamaa I, Shorohova E, Kushnevskaya H, Jalonen J (2017) Resilience of understory vegetation after variable retention felling in boreal Norway spruce forests: a ten-year perspective. For Ecol Manage 393: 12–28. https://doi.org/10.1016/j.foreco.2017.02.040.
Wikars LO (1995) Clear-cutting before burning prevents establishment of the fire-adapted Agonum quadripunctatum (Coleoptera: Carabidae). Ann Zool Fennici 32: 375–384. https://www.jstor.org/stable/23735543.
Wikberg S, Perhans K, Kindstrand C, Boberg Djupström L, Boman M, Mattsson L, Schroeder LM, Gustafsson L (2009) Cost-effectiveness of conservation strategies implemented in boreal forests: the area selection process. Biol Conserv 142: 614–624. https://doi.org/10.1016/j.biocon.2008.11.014.
Ylisirniö A-L, Hallikainen V (2018) Retention patches maintain diversity of epiphytic and epixylic indicator lichens more effectively than solitary trees. Scand J For Res 33: 320–331. https://doi.org/10.1080/02827581.2017.1415370.
143 viitettä.