Jaana Luoranen  ,
Minna Kivimäenpää,
Hanna Ruhanen,
Katri Himanen,
Johanna Riikonen
,
Minna Kivimäenpää,
Hanna Ruhanen,
Katri Himanen,
Johanna Riikonen
Douglaskuusen taimien pakkasenkestävyys on mäntyä ja kuusta huonompi: lyhytpäiväkäsittely douglaskuusen pakkasenkestävyyden parantajana
Luoranen J., Kivimäenpää M., Ruhanen H., Himanen K., Riikonen J. (2024). Douglaskuusen taimien pakkasenkestävyys on mäntyä ja kuusta huonompi: lyhytpäiväkäsittely douglaskuusen pakkasenkestävyyden parantajana. Metsätieteen aikakauskirja vuosikerta 2024 artikkeli 23013. https://doi.org/10.14214/ma.23013
Tiivistelmä
Ilmaston lämmetessä Suomessa aiemmin harvinaisten, eteläisempää alkuperää olevien puulajien viljely on herättänyt kiinnostusta. Lauhkealla ilmastovyöhykkeellä kasvavaa douglaskuusta (Pseudotsuga menziesii (Mirb.) Franco) on pidetty yhtenä vaihtoehtona metsäkuuselle sen kaarnakuoriaiskestävyyden vuoksi. Douglaskuusi on arka syyshalloille sekä pakkasvaurioille taimitarhakasvatuksen ja taimien pakkasvarastoinnin aikana. Yhtenä vaihtoehtona pakkasenkestävyyden parantamiseksi on taimituotannossa käytettävä lyhytpäivä- (LP) käsittely. Luonnonvarakeskuksen tutkimustaimitarhalla verrattiin ensimmäisen vuoden douglaskuusen taimien pakkasenkestävyyttä kuuseen (Picea abies (L.) H. Karst.) ja mäntyyn (Pinus sylvestris L.). Lisäksi testattiin kolmen viikon LP-käsittelyn vaikutuksia toisen vuoden taimien pakkasenkestävyyteen neljällä Suomesta kerätyllä siemenalkuperällä. Pakkasenkestävyydet määritettiin kontrolloiduilla pakkastesteillä kahtena syksynä sekä silmun puhkeamisvaiheessa. Douglaskuusi oli selvästi herkempi sekä syksyn pakkasille että keväthalloille kuin kotoperäiset havupuut. Douglaskuusen alkuperien välillä oli selviä eroja sekä niiden karaistumisrytmissä että reagoinnissa LP-käsittelyyn. LP-käsittely paransi silmujen, rangan ja neulasten pakkasenkestävyyttä syyskuun alussa kaikilla alkuperillä. Syyskuun ja lokakuun lopulla rangan ja neulasten pakkasenkestävyys parani käsittelyn seurauksena, mutta silmujen ei. Ilman käsittelyä rangan pakkasenkestävyys vaihteli –17 ja –25 °C:n välillä, ja vain Kouvolassa sijaitsevasta metsiköstä kerätty alkuperä saavutti pakkasvarastointiin vaadittavan kestävyyden. Tulosten perusteella douglaskuusen taimien karaistuminen on kotimaisia pääpuulajeja heikompaa. LP-käsittelyn avulla voidaan parantaa taimien alkusyksyn, ja jonkin verran myös loppusyksyn pakkasenkestävyyttä. Jos vieraslaji douglaskuusta halutaan viljelyllä Suomessa nykyistä laajemmin, on alkuperien valintaan ja niiden pakkasenkestävyyteen kiinnitettävä erityistä huomiota ja LP-käsittelyä kannattaa käyttää taimikasvatusvaiheessa parantamaan taimien kestävyyttä.
Avainsanat
Pinus sylvestris;
paakkutaimi;
Picea abies;
karaistuminen;
Pseudotsuga menziesii;
taimikasvatus
-
Luoranen,
Luonnonvarakeskus, Luonnonvarat, Suonenjoki
 https://orcid.org/0000-0002-6970-2030
Sähköposti
jaana.luoranen@luke.fi
https://orcid.org/0000-0002-6970-2030
Sähköposti
jaana.luoranen@luke.fi
-
Kivimäenpää,
Luonnonvarakeskus, Luonnonvarat, Suonenjoki
https://orcid.org/0000-0003-0500-445X
Sähköposti
minna.kivimaenpaa@luke.fi
-
Ruhanen,
Luonnonvarakeskus, Tutkimusinfrastruktuuripalvelut, Suonenjoki
https://orcid.org/0000-0001-8259-6570
Sähköposti
hanna.ruhanen@luke.fi
-
Himanen,
Luonnonvarakeskus, Tuotantojärjestelmät, Suonenjoki
https://orcid.org/0000-0002-6368-4636
Sähköposti
katri.himanen@luke.fi
-
Riikonen,
Luonnonvarakeskus, Tuotantojärjestelmät, Suonenjoki
https://orcid.org/0000-0001-7407-5759
Sähköposti
johanna.riikonen@luke.fi
Vastaanotettu 4.12.2023 Hyväksytty 16.4.2024 Julkaistu 16.5.2024
Katselukerrat 48937
Saatavilla https://doi.org/10.14214/ma.23013 | Lataa PDF

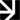
Supplementary Files
1 Johdanto
Vuotuiset keskilämpötilat Suomessa ovat kohonneet 1800-luvulta lähtien 2,3 °C (Mikkonen ym. 2015) ja ilmasto lämpenee tällä hetkellä noin puoli astetta vuosikymmenessä ilmastonmuutoksen takia (Lehtonen ym. 2020). Boreaalisella vyöhykkeellä kasvukauden ennustetaan pitenevän ja termisen talven lyhenevän sekä alku- että loppupäästä noin kahdella viikolla (Ruosteenoja ym. 2020). Kuusta (Picea abies (L.) H. Karst.) viljellään Suomessa paljon: 2020-luvun ensimmäisinä vuosina kuusen osuus istutetuista puun taimista on ollut noin 70 % (Suomen virallinen tilasto 2022). Kuusen on kuitenkin todettu olevan taloudellisesti tärkeistä puulajeistamme herkin kuivuudelle (Jansons ym. 2016) ja kuusen kannalta tuhonaiheuttajien on ennustettu hyötyvän ilmaston lämpenemisestä ja lisäävän siten kuusen tuhoriskejä (Venäläinen ym. 2020). Puulajirakenteen monipuolistamiseksi ja metsien hiilensidonnan turvaamiseksi on pohdittu vaihtoehtoja kuuselle (Ruotsalainen ym. 2022). Ilmaston ennustettu lämpeneminen mahdollistaa paitsi eteläisimpien kotimaisten, myös eräiden ei-kotoperäisten puulajien viljelyn tai sen laajentamisen. Toisaalta vieraslajeihin liittyy metsäluonnon monimuotoisuuden turvaamiseen liittyviä riskejä (Bärmann ym. 2023). Lisäksi metsäsertifiointijärjestelmät suhtautuvat ei-kotoperäisten lajien käyttöön kriittisesti (Viiri 2020).
Yhdeksi vaihtoehtoiseksi lajiksi esitetyn douglaskuusen (Pseudotsuga menziesii (Mirb.) Franco) (Ruotsalainen ym. 2022) on todettu kestävän kesäkuivuutta kuusta ja mäntyä (Pinus sylvestris L.) paremmin, joskin sopeutuneisuus kuivuuteen riippuu siementen alkuperästä (Nicolescu 2019). Kiinnostus douglaskuusen käyttöön metsänviljelyssä on lisääntynyt hieman viime vuosina (Suomen virallinen tilasto 2022). Viimeisten viiden vuoden aikana on istutettu 9000–47000 douglaskuusen taimea vuosittain, joka on ollut keskimäärin 0,02 % metsänviljelyyn toimitettujen taimien kokonaismäärästä (Suomen virallinen tilasto 2022). Ilmastonmuutos ja kuivuus lisäävät myös kuusia uhkaavan kirjanpainajan (Ips typographus L.) tuhojen riskiä Suomessa (Venäläinen ym. 2020). Kirjanpainaja, kuten jotkin muut Euroopassa tavattavat kaarnakuoriaislajit, voivat iskeytyä myös douglaskuuseen, mutta eivät ole aiheuttaneet sillä vakavaa tuhoa (EPPO Platform of PRAs 2018; Roques ym. 2019).
Douglaskuusella on kaksi alalajia, mereinen (P. menziesii var. menziesii) ja mantereinen (P. menziesii var. glauca (Beissn.) Franco). Näistä mereinen kasvaa nopeammin ja kookkaammaksi, mutta mantereinen kestää pakkasta paremmin (Nicolescu 2019). Länsi-Euroopan humidissa ilmastossa lauhkealla vyöhykkeellä douglaskuusen viljely lisääntyi toisen maailmansodan jälkeen, sen ollessa siellä tällä hetkellä toiseksi yleisin vieraspuulaji (van Loo ja Dobrowolska 2019).
Luontaisessa ympäristössään Pohjois-Amerikassa douglaskuusen merkittävin abioottinen tuhonaiheuttaja on keskitalven ja kevään alhaiset lämpötilat etenkin taimitarhoilla ja nuorissa taimikoissa (Nicolescu 2019). Bigrasin ym. (2001) kokoaminen tietojen mukaan douglaskuusen taimet voivat saavuttaa enintään –25 °C kestävyyden ja kasvukauden aikana pakkasenkestävyyttä ei juurikaan ole. Vastaavasti Suomessa kotoperäiset lajit voivat kestää –80 – –100 °C ja kasvukaudella kestävyys on eri tutkimuksissa vaihdellut 0ja –7 °C:n välillä (Bigras ym. 2001 ja viitteet taulukossa 1, s. 59). Samoin Nicolescu (2019) toteaa, että kahden ensimmäisen kasvukautensa aikana douglaskuusen taimet eivät kestä alle –25 °C:n lämpötiloja keskitalvella, mikä on selkeä riskitekijä nuorille taimille Suomessa.
Suomessa douglaskuusen huono taimivaiheen pakkasenkestävyys on todettu perustettaessa koeviljelmiä eri puolille maata 1930–1940-luvuilla (Silander ym. 2000) sekä viime vuosina pienessä mitassa tehdyissä taimien kasvatuskokeiluissa taimitarhoilla. Ruotsissa douglaskuusen istutustaimilla on havaittu runsaasti talvi- ja keväthallojen aiheuttamia tuhoja (Malmqvist ym. 2018). Taimitarhoilla ongelmia aiheuttaa myös taimien pakkasvarastoinnin aloittamisajankohdan määrittämisen vaikeus kaikilla havupuulajeilla, mutta erityisesti lajeilla, joiden kestävyydestä on vähän kokemusta. Tarkempia mittauksia douglaskuusen taimien karaistumisrytmistä tai syksyn ja kevään silmunpuhkeamisvaiheen pakkasenkestävyyseroista kotoperäisiin puulajeihin verrattuna ei myöskään ole.
Taimia voidaan taimitarhalla suojella pakkasvaurioilta ns. hallakastelulla. Sen tarpeen arvioinnissa taimien pakkasenkestävyyden rajana voidaan pitää –10 °C:n kestävyyttä. Tämä perustuu siihen, että hallakastelulla voidaan yleensä suojata taimia noin –7 …–8 °C:n lämpötiloilta (Rikala 2012). Lisäksi ilman lämpötilan on oltava 1–2 °C taimien pakkasenkestävyyden tasoa korkeampi hallakastelua aloitettaessa (Luoranen ja Rikala 2016). Taimet varastoidaan tavallisimmin talven yli pakkasvarastoissa –3 …–4 °C:n lämpötilassa.
Kestääkseen useamman kuukauden pituisen pakkasvarastoinnin, havupuun taimien on oltava riittävän karaistuneita varastoinnin alkaessa. Juurten kestävyys on versoa heikompi (Bigras ym. 2001). Versojen pakkasenkestävyyden määrittäminen on huomattavasti helpompaa ja siihen on nopeampia menetelmiä kuin juurille, minkä vuoksi taimien pakkasvarastointikestävyyden määrittämiseen on kehitetty versojen kestävyyteen perustuvia menetelmiä. Pohjois-Amerikassa havupuun taimien pakkasvarastointikestävyyden rajana on –18 °C (Ritchie ym. 2010). Ruotsalaistutkimuksissa kuusen ja männyn taimien määriteltiin olevan valmiita pakkasvarastointiin, kun taimien latvakasvaimet kestivät –25 °C (Lindström ym. 2014). Toisessa ruotsalaistutkimuksessa douglaskuusella versojen –20 °C:n kestävyyden on todettu takaavan riittävän pakkasvarastoinnin kestävyyden ainakin neljäksi kuukaudeksi, joskin alkuperien välillä on ollut selviä eroja (Malmqvist ym. 2017). Ruotsalaistutkimusten pakkasvarastoinnin kestävyyden määrittävät lämpötilat perustuvat taimen latvoista mitattujen altistusten jälkeisiin suhteellisiin ionivuotoihin, jotka kertovat solutasolla tapahtuvista vaurioista.
Taimien karaistuminen edellyttää, että taimien pituuskasvu on päättynyt ja silmun kehittyminen alkanut (Weiser 1970). Yksi- ja kaksivuotiaiden havupuuntaimien pituuskasvu päättyy syyskesällä yön pituuden ylittäessä lajille ja alkuperälle tyypillisen kriittisen arvon (Partanen 2004). Tätä ilmiötä hyödynnetään taimitarhoilla laajalti käytössä olevassa lyhytpäivä- (LP) käsittelyssä, jossa pimennysverhoilla keinotekoisesti pidennetään yötä niin, että taimien pituuskasvun päättyminen ja karaistuminen alkavat normaalia aiemmin (Rikala 2012). Suomessa menetelmää käytetään etenkin kuusen taimien kasvatuksessa. Pohjois-Amerikassa LP-käsittelyä on tutkittu ja käytetty myös douglaskuusella (esim. McCreary ym. 1978; Van Steenis 1992; MacDonald ja Owens 2010). Yksi keino myös Suomessa kasvatettavien douglaskuusen taimien talven- ja pakkasvarastoinnin kestävyyden varmistamiseksi voi olla LP-käsittely.
Tutkimuksen tavoitteena oli selvittää, (i) miten douglaskuusen eri alkuperien taimien karaistuminen etenee keskisen Suomen olosuhteissa verrattuna kuusen ja männyn taimiin, (ii) onko puulajien välillä eroa silmunpuhkeamisvaiheen pakkasenkestävyydessä sekä (iii) kuinka paljon taimitarhalla kesällä tehtävä LP-käsittely vaikuttaa douglaskuusen eri alkuperien taimien pakkasenkestävyyteen syksyllä. Hypoteesinamme oli, että douglaskuusi karaistuu syksyllä kuusta ja mäntyä hitaammin, erot ovat suurimmillaan silmunpuhkeamisvaiheessa keväällä, douglaskuusen alkuperien välillä on eroja pakkasenkestävyydessä, ja että lyhytpäiväkäsittely on tehokas keino parantaa douglaskuusen taimien pakkasenkestävyyttä syksyllä ja varmistaa pakkasvarastoinnin kestävyys kaikilla käytössä olleilla alkuperillä.
2 Aineisto ja menetelmät
2.1 Taimimateriaalin kasvatus
2.1.1 Yleistä taimikasvatuksesta
Tutkimus toteutettiin Luonnonvarakeskuksen (Luke) Suonenjoen tutkimustaimitarhalla (62°39´N, 27°03´E, 142 m meren pinnan yläpuolella). Tutkimus oli kaksiosainen; ensimmäisessä puulajien vertailukokeessa selvitettiin yksivuotiaiden kuusen, männyn ja douglaskuusen taimien eroja karaistumisessa syksyllä sekä silmun puhkeamisvaiheessa taimitarhalla, ja toisessa tutkittiin LP-käsittelyn vaikutusta syksyn karaistumiskehitykseen douglaskuusen taimilla. Molemmissa kokeissa kaikki taimet kasvatettiin PL81F (Plantek PL81F, 81 tainta/kennosto, yhden kennon tilavuus 85 cm3, kasvatustiheys 546 taimea m–2; BCC, Landskrona, Ruotsi) kennostoissa kohokasvatuksessa. Kennot oli täytetty peruslannoitetulla (1,0 kg m–3 liukoista lannoitetta, jossa NPK 16-4-7 kg m–3 + hivenravinteita sekä 6,5 % hidasliukoista typpeä) ja kalkitulla (1,8 kg m–3) vaalealla rahkaturpeella (Kekkilä White 420 W F6; Kekkilä Professional Co., Vantaa).
2.1.2 Puulajien vertailukokeen taimet
Puulajien vertailukokeessa douglaskuusen Mustilan arboretumista ostettua, Kouvolan metsikkökerättyä alkuperää (K09-21-475; tarkempi mantereinen alkuperä Kanadassa tuntematon, mutta sen oletetaan olevan Mustilan vanhimman, vuonna 1910 istutetun metsikön jälkeläisiä, jonka alkuperä on Quesnel lähellä Prince Georgea (suullinen tiedonanto, J. Reinikainen)) olevat siemenet esikäsiteltiin Suonenjoen siemenlaboratoriossa. Esikäsittelyssä siemeniä liotettiin ilmastetussa vedessä 16 tunnin ajan 15 °C:ssa, minkä jälkeen kelluvat ja astian pohjalle laskeutuneet siemenet erotettiin toisistaan. Röntgenkuvauksen perusteella kelluvat siemenet todettiin heikkolaatuisiksi ja tämä osite hylättiin. Pohjaositteen siemenet kylvettiin käsin 20.4.2022 (yksi siemen/kenno) pintakuivauksen jälkeen. Kuusen (alustavasti testattu SV171; EY/FIN M29-19-0033; käyttöalue eteläisessä ja keskisessä Suomessa 1110–1310 d.d., leveysasteet 60–62 (osin 63)° (Ruokavirasto 2024)) siemenet kylvettiin koneellisesti 3.5.2022 ja männyn (testattu SV323; EY/FIN M29-15-0012; soveltuu käytettäväksi keskisessä Suomessa, leveysasteet 61–63° (Ruokavirasto 2024)) 15.5.2022. Kuusen ja männyn siemenet peitettiin kylvön jälkeen ohuella hiekkakerroksella. Kylvön jälkeen douglaskuuset olivat aluksi lämmitetyssä kasvihuoneessa, josta ne siirrettiin 13.5.2022 tuotantomittakaavan taimitarhakasvatukseen muovihuoneeseen. Kuusen ja douglaskuusen taimet olivat muovihuoneessa koko kokeen ajan, männyn taimet siirrettiin ulkokasvatuskentälle 12.7.2022. Muovihuoneen ovia pidettiin auki syksyllä kovimpia pakkasöitä lukuun ottamatta. Kasvatuksen aikana taimia kasteltiin säännöllisesti ja lannoitettiin Metsä Superex (NPK 22-5-16, Kekkilä Professional Co., Vantaa) lannoitteella. Kuusia ja douglaskuusia kasteltiin ja lannoitettiin samalla ohjelmalla. Lannoituskertoja oli yhteensä 13 välillä 8.6.–15.9.2022 ja taimet saivat yhteensä 24,4 g m–2 typpeä, 5,7 g m–2 fosforia ja 17,7 g m–2 kaliumia. Männyn taimia lannoitettiin kuusi kertaa 8.6.–22.7.2022 välillä ja ne saivat 10,2 g m–2 typpeä, 2,4 g m–2 fosforia ja 7,4 g m–2 kaliumia. Kunkin puulajien taimia oli kasvatuksessa yhteensä 1400 tainta.
Puulajien vertailukokeessa kaikkien puulajien taimet pakattiin pahvilaatikoihin marraskuun alussa 2022 ja laatikot siirrettiin pakkasvarastoon –3 °C:seen käytettäväksi talvella 2023 silmunpuhkeamisvaiheen pakkasenkestävyyden tutkimiseen. Pakkasvarastosta taimet otettiin sulamaan +5 °C:seen, ja viiden vuorokauden jälkeen taimet siirrettiin helmikuun alussa 2023 säädeltyihin kasvatuskaappeihin (Conviron Adaptis A1000, Conviron, Winnipeg, Kanada), jossa oli 18 tunnin päivä ja 6 tunnin yö ja päivällä 20 °C ja yöllä 18 °C. Kun taimien silmut olivat silmin havaittavasti turvonneet, niille tehtiin pakkastestit. Silmujen turpoamiseen vaaditut lämpösummat (+5 °C kynnysarvolla) olivat douglaskuuselle 101,5 d.d. (13.–20.2.2023), kuuselle 116 d.d. (6.–14.2.2023) ja männylle 72,5 d.d. (31.1.–5.2.2023). Kasvatuksen aikana taimien juuripaakut pidettiin koko ajan kosteina.
2.1.3 Lyhytpäiväkäsittelykokeen taimet
LP-käsittelyn vaikutuksia tutkivassa kokeessa käytettiin kolmea mantereista douglaskuusen siemenalkuperää, jotka oli kerätty Punkaharjun Laukansaaresta kasvaneista metsiköistä (taulukko 1). Lisäksi mukaan otettiin Mustilan arboretumista ostettua, Kouvolassa kasvaneesta douglaskuusimetsiköstä kerättyä siementä. Kouvolan alkuperää olevat siemenet kylvettiin suoraan PL81F-kennoihin lämmittämättömään muovihuoneeseen 19.5.2020. Muiden alkuperien siemenet hajakylvettiin turpeella täytetyille alustoille lämmitettyyn kasvihuoneeseen 15.5.2020. Orastuminen oli näissä iäkkäissä siemenerissä varsin epätasaista ja jatkui pitkään, joten sirkkataimien koulintaa jouduttiin tekemään 3.–18.6. sitä mukaa, kun uusia sirkkataimia ilmestyi. Sirkkataimet koulittiin turpeella täytettyihin PL81F-kennostoihin, yksi taimi kuhunkin paakkuun. Muutaman päivän juurtumisajan jälkeen kennostot siirrettiin muovihuoneeseen Kouvolan alkuperän viereen. Kaikki taimet siirrettiin muovihuoneesta ulkokasvatuskentälle 15.8.2020. Syyskuun lopulla oli pakkasöitä, joiden aikana taimia suojattiin harsoilla. Lokakuun alussa taimet siirrettiin takaisin muovihuoneeseen, jossa ne olivat paremmin suojassa yöpakkasilta. Muovihuoneen ovia pidettiin päivisin ja lämpiminä öinä auki.
| Taulukko 1. Lyhytpäiväkäsittelyn vaikutuksia tutkivassa kokeessa käytettyjen douglaskuusen alkuperistä käytetyt nimet, siementen keräysmetsikön tunnukset, siemenerän tunnus sekä keräysmetsikön perimän mantereinen alkuperä Kanadassa. Kaikki siemenerät olivat vapaapölytteisiä. | ||||
| Alkuperä | Keräysmetsikkö | Siemenerän tunnus | Tuleentumisvuosi | Keräysmetsikön perimän alkuperä |
| Larch Hills | Mv 126 | EU/FIN G04-89-0078 | 1989 | Brittiläinen Kolumbia |
| Salmon River | Mv 123 | EY/FIN G4-89-77 | 1989 | Brittiläinen Kolumbia, 50°N, 126°W, 660 m |
| Valemount | Mv 125/2 | EY/FIN G04-89-0079 | 1989 | Brittiläinen Kolumbia |
| Kouvola | K09-19-631A | 2019 | tuntematon | |
Taimikennostot (37 kappaletta, 10 kennoa/alkuperä, paitsi Valemountin alkuperällä vain 7) olivat kohokasvatuksessa ja niitä kasteltiin ja lannoitettiin tarpeen mukaan. Taimet käsiteltiin Proline 250 EC (protiokonatsoli 250 g l–1, 25 % (w/v)) kasvinsuojeluaineella harmaahometta vastaan 4.11.2020. Sen jälkeen taimet pakattiin pahvilaatikoihin, yksi kennosto/laatikko, ja siirrettiin talveksi pakkasvarastoon (–3 °C) marraskuun 2020 alussa. Taimet siirrettiin 11.5.2021 +8–9 °C:n lämpötilaan sulamaan ja 17.5.2021 ulos kasvatuskentälle, jossa niitä kasvatettiin LP-käsittelyjen alkamiseen saakka. Toukokuussa 2021 inventoitiin kasvatuskelpoisten taimien määrät, jotka olivat seuraavat: Kouvola 69 %, Larch Hills and Salmon River molemmilla 93 % ja Valemount 83 %. Kasvukauden 2021 aikana taimia kasteltiin tarpeen mukaan ja lannoitettiin seitsemän kertaa (Metsä Superex NPK 22-5-16, Kekkilä Professional Co., Vantaa). Kasvukauden 2021 aikana taimet saivat yhteensä 15,8 g m–2 typpeä, 3,6 g m–2 fosforia ja 11,5 g m–2 kaliumia. Heinäkuussa taimikennostot jaettiin kahteen osaan, joista viisi kennostoa/alkuperä (Valemount vain kolme) LP-käsiteltiin kolme viikkoa alkaen 6.7.2021 käyttäen pimennysverhoa 12 h ajan (klo 19:00–07:00). Loput kennostot (viisi kennostoa/alkuperä, Valemount 4 kennostoa) jätettiin käsittelemättömäksi kontrolliksi.
2.2 Sääolosuhteet
Suonenjoen ilmastoa kuvaavat Ilmatieteenlaitoksen pitkän ajan (1911–2020) säätilastot ovat peräisin Kuopion Maaningan sääasemalta (69°77´N, 19°90´E; Ilmatieteenlaitos, vertailukausi). Kylmin kuukausi on helmikuu, jolloin keskilämpötila on –8,4 °C. Pakkaspäiviä (lämpötila T < 0 °C) on keskimäärin 169 ja kylmiä päivä (T < –10 °C) 57 vuodessa. Lämpimin kuukausi on heinäkuu (17,1 °C). Vuotuinen sademäärä on keskimäärin 617 mm. Lunta on vertailukaudella ollut marraskuusta (mediaani lumen syvyys 15.11. 0cm ja 31.11. 4 cm) huhtikuuhun (mediaani 15.4. 12 cm) ja runsaslumisin kuukausi on maaliskuu (47 cm). Ilmatieteen laitoksen 1 × 1 km hilapisteaineistosta laskettuna Suonenjoen viiden vuoden (2016–2021) kasvukauden lämpösumma on ollut keskimäärin 1384 d.d.
Luken Suonenjoen tutkimustaimitarhan sääaseman mittausten mukaan vuonna 2021 kesä- ja heinäkuu olivat pitkän ajan (2000–2022) keskiarvoihin verrattuna selvästi lämpimämpiä ja vähäsateisempia (taulukko 2). Touko-, syys- ja lokakuu 2021 olivat puolestaan viileämpiä ja elokuu runsassateinen verrattuna keskimääräisiin arvoihin Suonenjoella vuoden 2000 jälkeen. Heinäkuu 2022 oli lämpötiloiltaan keskimääräinen, touko- ja syyskuu keskimääräistä viileämpiä ja kesä-, elo- ja lokakuu lämpimämpiä. Koko kasvukausi 2022 oli keskimääräistä vähäsateisempi. Muovihuoneessa sekä keski- että alimmat lämpötilat olivat 5–10 °C ulkona mitattuja korkeammat.
| Taulukko 2. Kuukausittaiset keskilämpötilat (ka.; °C; 2 m korkeudelta mitattuna) ja alimmat lämpötilat (10 cm korkeudelta maanpinnasta mitattuna) sekä sadesummat (mm) kasvukausien 2021–2022 aikana sekä pitkänajan kuukausittaiset keski- ja alimmat lämpötilat sekä sadesummat vuosilta 2000–2022 Luonnonvarakeskuksen Suonenjoen koetoiminta-aseman sääasemalla. Kasvukauden 2022 osalta suluissa olevat lukemat ovat muovihuoneessa mitattuja keski- ja alimpia lämpötiloja kasvuston yläpuolella. | |||||||||
| 2021 | 2022 | 2000–2021 | |||||||
| Lämpötila | Sadesumma | Lämpötila | Sadesumma | Lämpötila | Sadesumma | ||||
| Kuukausi | ka. | alin | ka. | alin | ka. | alin | |||
| toukokuu | 7,5 | –8,2 | 49,3 | 8,7 (17,4) | –6,1 (7,0) | 11,8 | 9,5 | –8,8 | 44,6 |
| kesäkuu | 18,6 | 1,9 | 40,4 | 16,0 (21,4) | 1,4(11,5) | 47,9 | 14,4 | –3,9 | 64,0 |
| heinäkuu | 20,0 | 4,1 | 54,2 | 17,2 (22,0) | 5,6 (12,8) | 67,6 | 17,3 | –0,7 | 77,6 |
| elokuu | 14,2 | 3,0 | 140,5 | 17,1 (22,1) | 4,0 (11,6) | 41,2 | 15,2 | –1,9 | 70,0 |
| syyskuu | 7,3 | –5,4 | 40,0 | 7,8 (12,9) | –4,5 (2,7) | 29,7 | 9,8 | –7,4 | 55,7 |
| lokakuu | 5,8 | –9,0 | 80,6 | 5,1 (7,6) | –8,5 (–1,4) | 21,5 | 3,8 | –17,2 | 50,3 |
2.3 Pakkasenkestävyyden määrittäminen
Puulajien vertailukokeessa taimien pakkasenkestävyys määritettiin kaksi kertaa syksyllä 2022 (19.9. ja 24.10.) ja kerran lyhyen kasvatuskaappikasvatuksen jälkeen talvella 2023, kun silmut olivat hieman turvonneita. LP-kokeen taimet altistettiin kylmälle kolmena ajankohtana syksyllä 2021 (8.9., 28.9. ja 20.10.). Puulajien vertailukokeessa jokaiselta kokeessa olleelta kolmelta suuralustalta valittiin satunnaisesti 96 tainta, yhteensä 288 tainta kutakin puulajia muutama vuorokausi ennen syksyn 2022 pakkasaltistuksia. Kunkin puulajin taimet laitettiin pienemmiksi sahattuihin PL81F kennostoihin, 36 tainta (12 jokaiselta suuralustalta) altistettavaksi kahdeksaan testilämpötilaan eri testikerroilla. Yhden puulajin taimia oli siis kahdeksassa halkaistussa kennostossa kaikilla testikerroilla. Talvella 2023 eri puulajien testit tehtiin eri aikoina johtuen puulajien erilaisista haluttuun kehitysvaiheeseen vaadittavista lämpösummista.
LP-kokeessa taimien pakkasenkestävyyden määrittämistä varten kunkin alkuperän molempien käsittelyiden taimet jaettiin satunnaisesti taimikennostoihin niin, että yhdessä kennostossa oli 10 tainta jokaisesta alkuperästä ja käsittelystä. Taimikennostoja oli siten yhteensä 23 kappaletta. Taimien vähäisyyden takia toisella testikerralla 28.9. ei ollut mukana Valemountin alkuperää ja testilämpötiloja oli vain seitsemän, kun niitä muilla testikerroilla oli kahdeksan.
Pakkasenkestävyyden määrittämiseen käytettiin neljää ilmajäähdytteistä olosuhdekaappia (WT600/70, Weiss Umweltechnik GmbH, Reiskirchen-Lidenstruth, Saksa). Jokainen kaappi ohjelmoitiin saavuttamaan yksi tavoitelämpötila. Testit tehtiin riittävän lämpötilamäärän saamiseksi kahtena peräkkäisenä yönä. Puulajikokeessa syksyllä 2022 kaappien lämpötila vastasi taimien näytteenottohetken ulkolämpötilaa testin alussa ja silmunpuhkeamisvaiheessa keväällä 2023 kasvatuskaappien päivälämpötilaa. LP-kokeessa kaappien lämpötila oli testin alussa +5 °C. Molemmissa kokeissa lämpötila laskettiin aluksi 3 °C tunnissa –5 °C:seen, jossa lämpötila pidettiin kolme tuntia. Sen jälkeen lämpötila laskettiin 5 °C tunnissa tavoitelämpötilaan, jossa taimia pidettiin kolme tuntia ja sen jälkeen lämpötilaa nostettiin 5 °C tunnissa +5 °C:seen, jossa taimia pidettiin 1–5 vuorokautta (aika riippui altistuslämpötilasta) juuripaakkujen sulattamiseksi. Kontrollilämpötilana oli +5 °C. Kunkin testikerran lämpötilat on esitetty taulukossa 3. Käytössä oli neljä olosuhdekaappia, joten testejä tehtiin kahtena tai kolmena peräkkäisenä päivänä.
| Taulukko 3. Syksyllä 2021 (LP-koe; kursivoitu teksti) ja 2022 (puulajikoe) sekä talvella 2023 (puulajikoe, silmunpuhkeamisvaihe) toteutettujen pakkastestien ajankohdat, douglaskuusen alkuperät (LP-koe vuonna 2021) ja puulajit (vuoden 2022 koe) kullakin testikerralla sekä testilämpötilat. | ||
| Ajankohta | Alkuperät tai puulaji | Testilämpötilat, °C |
| 8.9.2021 | Kouvola, Larch Hills, Salmon River, Valemount | +5, –2, –4, –6, –8, –10, –12, –15 |
| 29.9.2021 | Kouvola, Larch Hills, Salmon River | +5, –5, –7, –10, –12, –20, –25 |
| 20.10.2021 | Kouvola, Larch Hills, Salmon River, Valemount | +5, –8, –10, –15, –20, –23, –25, –30 |
| 19.9.2022 | Kuusi | +5, –5, –8, –10, –12, –15, –18, –25 |
| 19.9.2022 | Mänty | +5, –8, –10, –12, –15, –18, –20, –25 |
| 19.9.2022 | Douglaskuusi | +5, –5, –8, –10, –12, –15, –18, –25 |
| 24.10.2022 | Kuusi | +5, –20, –25, –30, –35, –40, –50, –60 |
| 24.10.2022 | Mänty | +5, –10, –15, –20, –25, –30, –35, –45 |
| 24.10.2022 | Douglaskuusi | +5, –5, –10, –15, –20, –25, –30, –45 |
| 14.2.2023 | Kuusi | +5, –3, –6, –8, –10, –12, –15, –20 |
| 5.2.2023 | Mänty | +5, –3, –6, –8, –10, –12, –15, –20 |
| 20.2.2023 | Douglaskuusi | +5, –3, –6, –8, –10, –12, –15, –20 |
Pakkasaltistusten jälkeen taimet siirrettiin kennostoissaan kasvihuoneeseen, jonka lämpötila pidettiin +20 °C:ssa. Päivänpituutta jatkettiin suurpainenatriumlampuilla 18 tuntiin ja taimia kasteltiin säännöllisesti pitäen juuripaakut koko ajan kosteina. Syksyn testien taimista määritettiin pakkasvauriot kahden viikon kuluttua altistuksesta. Talven testeissä vauriot määritettiin kuuden (mänty), viiden (kuusi) ja seitsemän (douglaskuusi) viikon kuluttua, jotta kaikki puhkeamiskykyiset silmut olivat varmasti puhjenneet ja taimet kasvussa. Vauriot määritettiin erikseen halkaistuille silmuille ja rangalle sekä neulasille. Ruskettuneiden ja kuivien neulasten osuus arvioitiin silmävaraisesti 10 % luokissa. Halkaistusta rangasta arvioitiin myös prosentteina vaurioituneiden osien pituudet. Syksyllä silmut arvioitiin joko vaurioituneiksi (1) tai eläviksi (0). Halkaistu silmu ja rangan nila ja jälsi luokiteltiin vaurioituneeksi, jos niiden terveen vihreä väri oli muuttunut ruskeaksi tai normaalia vaaleammaksi verrattuna altistamattomiin +5 °C:n kontrollitaimiin. Silmunpuhkeamisvaiheen testissä silmu luokiteltiin eläväksi, jos se oli lähtenyt kasvuun, muutoin se oli vaurioitunut. Rangan ja silmujen arvioinnissa käytettiin apuna stereomikroskooppia.
Eri kasvinosille laskettiin kaksi estimaattia pakkasenkestävyydelle käyttäen yleistettyä epälineaarista sekamallia (NLMIXED; SAS for Windows 9.4, Usage Note 56992; SAS Institute Inc., Cary, NC, USA). Laskennan tarkempi kuvaus löytyy Riikosen ja Luorasen (2020) julkaisusta. Yleisesti pakkasenkestävyystutkimuksessa käytetyt DT50 arvot laskettiin lämpötilana, jossa 50 % neulasista, rangasta tai silmuista oli vaurioitunut. Käytännön taimitarhakasvatusta ajatellen lämpötilat, joissa puolet taimista on vaurioitunut, ovat liian alhaisia. Siksi laskimme myös DT05 arvot, jos se oli mahdollista. Nuo arvot kuvaavat lämpötiloja, joissa 5 % kasvinosasta tai taimista (silmut) oli vaurioitunut. Tämä vauriotaso valittiin, koska se vastaa taimierien myyntikelpoisuudelle asetettuja vaatimuksia.
Pakkasenkestävyysarvojen laskenta tehtiin LP-kokeessa yhden testikerran sisällä niin, että kaikki alkuperät ja käsittelyt olivat samassa mallissa. Mallin antamien DT50 -arvojen tilastolliset erot testattiin NLMIXED analyysin yhteydessä pareittain käyttäen delta-menetelmää. Tilastolliset erot alkuperien ja käsittelyiden välillä yhden testikerran sisällä laskettiin Waldin testillä. Näiden tilastollisten testien kuvaukset löytyvät Lapin ja Luorasen (2018) julkaisusta. Puulajien vertailukokeessa kunkin kasviosan pakkasenkestävyydet syksyn testiajankohdille laskettiin yhdellä mallilla. Silmunpuhkeamisvaiheen testit tehtiin erikseen kullekin puulajille. Puulajit olivat kasvatuksen ja altistuksen jälkeen erillään toisistaan ilman selkeää koejärjestelyä, joten puulajeja ei voitu verrata tilastollisesti toisiinsa. Analyyseissä rangalle ja neulasille käytettiin taimikohtaisia mitattuja numeerisia vaurio-osuuksia. Silmuille laskettiin suuralustoittain (puulajit) tai lohkoittain (LP-koe) vaurioituneiden päätesilmujen osuudet, joita käytettiin tilastollisissa testeissä.
LP-kokeessa DT05 arvojen laskentaa hankaloitti se, että etenkin viimeisellä testikerralla lokakuun lopulla myös kontrollilämpötilassa oli vaurioita (ks. liite L1 estimointikäyrät eri testikerroille). Laskimme DT05 arvot niille kerroille, joille se oli mahdollista (liite L2, kuva L2.1). Koska tavoitteemme oli vertailla eri alkuperiä ja LP-käsittelyn vaikutusta douglaskuusen kestävyyteen syksyn eri vaiheissa, esitämme tässä tulokset vain käyttäen DT50 estimointia, jonka tulokset kuvaavat eroja estimointikäyrien käännepisteessä ja taimien lievät vauriot ennen altistustestien tekemistä eivät häiritse alkuperien ja käsittelyiden vertailua merkittävästi.
Kaikissa analyyseissä erojen katsottiin olevan tilastollisesti merkitseviä, kun p-arvo oli <0,05.
3 Tulokset
3.1 Puulajien vertailukoe
Puulajien vertailukokeessa vauriokäyrien estimointi oli mahdollista, kun vauriohavaintoja oli sekä osittain että kokonaan vaurioituneista kasvinosista (kuva 1) eikä kontrollilämpötilana käytetyssä +5 °C lämpötilassa esiintynyt vaurioita. Siten sekä DT05 että DT50 arvojen estimointi oli mahdollista. Estimoinnin tuloksena saadut parametrien estimaatit on esitetty liitteessä L3.
Kuva 1. Havaitut (symbolit) ja estimoidut (viivat) a, d, g) vaurioituneiden neulasten, b, e, h) vaurioituneen rangan ja c, f, i) vaurioituneiden silmujen osuudet yksivuotiailla a–c) douglaskuusen, d–f) kuusen ja g–i) männyn taimilla määritettynä 19.–22.9.2022 ja 24.–26.10.2022 sekä keväisen silmujen puhkeamisen aikaan.
Syyskuun lopulla douglaskuusen kaikki kasvinosat kestivät vähintään –3 °C lämpötilan (DT05) vähäisin vaurioin (kuva 2). Vastaavasti kuusella DT05 arvot vaihtelivat –5 °C:n (neulaset) ja –7 °C:n (ranka) välillä. Männyllä neulaset kestivät vähäisin vaurioin vain –4 °C:n lämpötilan, mutta ranka ja silmut vähintään –7 °C. Lokakuun lopulla douglaskuusen eri osat kestivät vähäisin vaurioin noin –10 °C ja männyt –13 °C. Kuusella silmut kestivät –14 °C, mutta neulaset ja ranka jo vähintään –20 °C.
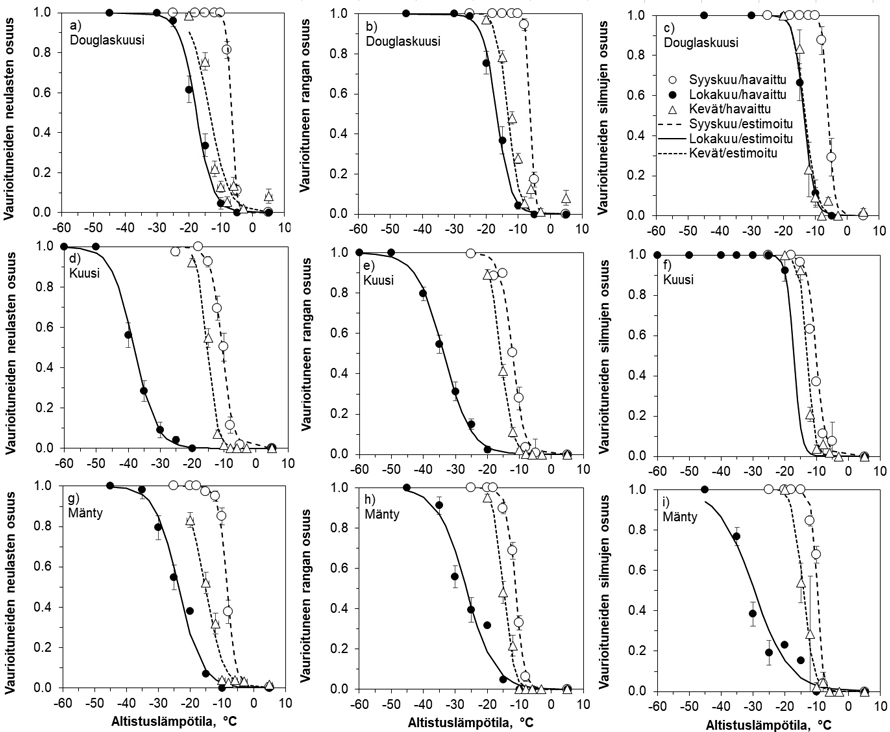
Kuva 2. Douglaskuusen, kuusen ja männyn taimien a) neulasten, b) rangan ja c) silmujen pakkasenkestävyydet syys- ja lokakuun lopulla 2022 sekä silmun puhkeamisvaiheessa lepovaiheen jälkeen. Pakkasenkestävyydelle laskettiin estimaatit sekä sille lämpötilalle, jossa neulasista, rangasta tai silmuista vaurioitui 5 (DT05) tai 50 % (DT50). Pystyviivat pylväiden päissä kuvaavat arvojen keskivirheitä. Vaakakatkoviivat kuvaavat hallan (–10 °C) ja pakkasvarastoinnin (–18 °C) kestävyysrajoja syksyllä.
DT50 arvojen mukaan syyskuun testissä taimien eri osien pakkasenkestävyydet vaihtelivat –6 ja –12 °C:n välillä (kuva 2). Douglaskuuset olivat vähiten kestäviä. Vähintään –10 °C:n kestävyyden olivat saavuttaneet männyn rangat ja silmut sekä kuusen kaikki osat. Lokakuun testissä douglaskuuset olivat edelleen vähiten kestäviä ja kaikkien osien kestävyys jäi alle –18 °C:seen. Silmut olivat altteimpia pakkasvaurioille. Kuusella rangat ja neulaset sekä männyllä rangat ja silmut kestivät –25 °C:n lämpötilat, mutta männyn neulasten kestävyys jäi hieman heikommaksi. Kuusen silmujen kestävyys jäi vain noin –18 °C:seen.
Silmujen puhkeamisvaiheessa douglaskuusi oli edelleen vähiten kestävä DT50-arvojen ollessa –12 ja –13 °C:n välillä kaikilla kasvinosilla, kuusen ja männyn rankojen ja neulasten kestettyä 2–4 °C alhaisempia lämpötiloja (kuva 2). Lieviä vaurioita aiheuttavien lämpötilojen (DT05) osalta douglaskuusen rangat ja neulaset kestivät vain enintään –5 °C ja silmut –8 °C, kun männyllä ja etenkin kuusella kestävyydet olivat 1–7 °C parempia (kuva 2).
3.2 Lyhytpäiväkäsittelykoe
DT50-arvojen perusteella sekä käsittelemättömillä että LP-käsitellyillä taimilla oli alkuperien välillä eroja niin neulasten, silmujen kuin rangan kestävyydessä kaikkina ajankohtina (p < 0,001 kaikkina kertoina kaikille kasvin osille ja molemmille käsittelyille; liite L4 taulukko L4.1). Pakkasenkestävyyden NLMIXED analyysin parametrien estimaatit sekä käsittelyiden ja alkuperien parittaiset vertailut kasvinosittain ja testikerroittain on esitetty liitteessä L4. LP-käsitellyt taimet kestivät vähintään –9 °C syyskuun alussa, kun käsittelemättömät taimet kestivät –5 – –7 °C, ja käsittelyiden ero kestävyydessä vaihteli alkuperästä ja kasvinosalta riippuen 4 ja 7 °C:n välillä (kuva 3a–c). Kestävimpiä olivat Kouvolan alkuperän taimet.
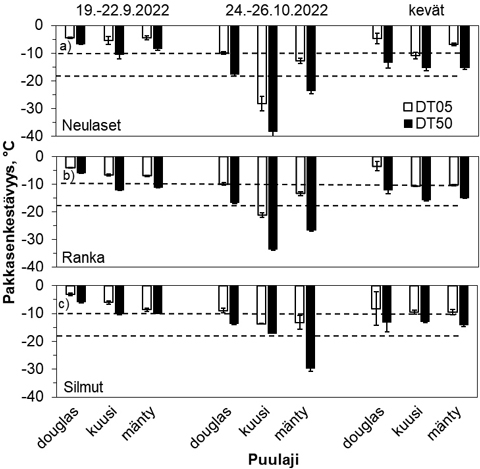
Kuva 3. Lyhytpäiväkäsiteltyjen ja käsittelemättömien kaksivuotiaiden douglaskuusen paakkutaimien eri alkuperää olevien a, d, g) neulasten, b, e, h) rangan ja c, f, i) silmujen pakkasenkestävyys (DT50) a–c) 8.9., d–f) 29.9. ja g–i) 20.10.2021 altistustesteissä Suonenjoen tutkimustaimitarhalla. Pienet kirjaimet pylväiden alla kertovat tilastolliset erot (p < 0,05) alkuperien välillä kontrollitaimilla ja isot kirjaimet LP-käsiteltyjen taimilla yhden testikerran ja kasvinosan sisällä. Tähtimerkki (*) kertoo tilastollisen eron käsittelyiden välillä yhden alkuperän sisällä. Pystyviivat pylväiden päissä kuvaavat DT50 arvojen keskivirheet. Vaakakatkoviivat kuvaavat kuvissa a–c –10 °C hallankestävyysrajaa, muissa kuvissa –18 °C:n ajateltua pakkasvarastoinnin kestävyysrajaa.
Syyskuun lopulla LP-käsittely paransi vain neulasten kestävyyttä noin 4 °C niin, että käsiteltyjen taimien neulaset kestivät vähintään –17 °C (kuva 3d). Silmujen ja rangan kestävyys oli noin –13 °C muutoin paitsi Kouvolan alkuperällä se oli jo vähintään –16 °C, ja silläkin käsittelemättömien taimien silmut kestivät vain –13 °C (kuva 3e–f).
Lokakuun 20. päivän testissä rangan ja neulasten osalta trendi oli, että LP-käsittely paransi kestävyyttä, joskin tilastollisesti merkitseviä erot käsittelyiden välillä olivat vain Larch Hillsin alkuperällä (kuva 3g–h). Se oli muutenkin pakkasenkestävyydeltään heikompi kuin etenkin Kouvolan alkuperän taimien neulaset ja ranka. Kouvolan alkuperän rangat kestivät ilman LP-käsittelyä vähintään –25 °C (kuva 3h), kun LP-käsittelyn jälkeen ranka kesti –33 °C (kuva 3h) ja neulaset jopa –37 °C (kuva 3g). LP-käsittely ei vaikuttanut silmujen kestävyyteen ja alkuperien välillä erot olivat pieniä (kuva 3i). Heikoin kestävyys oli Larch Hills alkuperän taimien silmuilla, jotka kestivät vain –14 °C (kuva 3i). LP-käsitellyillä taimilla muutosta silmujen kestävyydessä ei juurikaan ollut tapahtunut syyskuun lopulta ja enimmillään silmut kestivät –19 °C Kouvolan alkuperällä.
4 Tulosten tarkastelu
Douglaskuusen taimien pakkasenkestävyys oli lievästi heikompi alkusyksyllä verrattuna männyn ja etenkin kuusen taimien kestävyyteen, mutta loppusyksyllä sen kestävyys oli selvästi heikompaa kotoperäisiin havupuihin verrattuna. Silmujen puhkeamisvaiheessa etenkin rangan ja neulasten pakkasenkestävyys olivat douglaskuusella muita lajeja heikompia. Kaikilla tutkimuksessa mukana olleilla douglaskuusen alkuperillä taimien karaistuminen syksyllä oli suhteellisen hidasta, mutta LP-käsittelyllä sitä voitiin nopeuttaa jonkin verran.
Douglaskuusen heikompi pakkasenkestävyys kotoperäisiin havupuihin verrattuna oli odotettua. Alenevat lämpötilat ja lämpötilan vaihtelu ovat tärkeitä taimien karaistumista edistäviä tekijöitä havupuilla silmun muodostumisen jälkeen (Weiser 1970). Etenkin männyllä syksyn sääolosuhteet vaikuttavat karaistumiseen, sillä maksimikestävyyden saavuttaakseen se tarvitsee lähellä nollaa olevia lämpötiloja jo alkusyksyllä (Bigras ym. 2001; Repo ym. 2001). Kokeessamme männyn taimet siirrettiinkin ulos jo loppukesästä. Aiemmissa tutkimuksissa männyn neulaset ovat kestäneet lokakuun puolivälin jälkeen jo –30 °C (Di ym. 2019) eli tutkimuksessamme männyn neulaset kestivät hieman vähemmän, joskin riittävästi esimerkiksi pakkasvarastointia ajatellen. Kuusella taas asteittain aleneva lämpötila syksyllä on tehokkain parhaan kestävyyden saavuttamisessa (Bigras ym. 2001), eli kokeessamme taimien pitäminen muovihuoneessa ovet avoimena todennäköisesti varmisti kuuselle parhaan kestävyyden lokakuun testikertaan mennessä ainakin neulasille ja rangalle. Silmujen kestävyys olisi voinut parantua viileämmässä.
Vertailtaessa eri puulajien silmujen kestävyyksiä DT50 arvoina, männyllä silmut kestivät loppusyksystä pakkasta parhaiten. Sen sijaan douglaskuusen silmujen kestävyys oli heikointa vertailtaessa vähäisiä vaurioita kuvaavia DT05 arvoja, mutta kuusen ja männyn välillä ei ollut niissä eroja. Douglaskuusen pohjoisamerikkalainen alkuperä oli kuusen ja männyn pluspuihin verrattuna leveyspiiriltään 10–12° eteläisempi. Puulajien vertailukokeessa douglaskuusen pituuskasvu jatkui aina 10.9. saakka, kun kuusen taimet lopettivat kasvunsa jo 26.8. (Luoranen, julkaisematon). Näin ollen sen karaistuminen alkoi kuusta myöhemmin ja selittää osin sen muita puulajeja heikompaa kestävyyttä.
Eri vuosina tekemissämme kokeissa douglaskuusen taimien pakkasenkestävyydet erosivat toisistaan: puulajien vertailukokeessa taimet olivat syyskuun lopulla ja lokakuussa vähemmän kestäviä kuin LP-kokeessa. Eroa voisi selittää taimien ikä, sillä puulajien vertailukokeessa taimet olivat yksi- ja LP-kokeessa kaksivuotiaita. Kuusella tosin yksi- ja kaksivuotiaiden taimien karaistumisessa ei ole ollut eroja (Konttinen ym. 2007). Todennäköisin syy vuosien välisiin eroihin pakkasenkestävyydessä oli erilaiset kasvuolosuhteet. Puulajikokeessa douglaskuusen ja kuusen taimet olivat syksylläkin muovihuoneessa, kun LP-kokeen taimet olivat ulkona. Ulkolämpötiloiltaan vuodet olivat suhteellisen samanlaisia, mutta ovien aukipitämisestä huolimatta lämpötilat muovihuoneessa olivat syksyllä korkeammat sekä yö- ja päivälämpötilan vaihtelu vähäisempää. Koska douglaskuusten kestävyys oli tutkimuksemme mukaan heikko ensimmäisen kasvukauden lopulla, niiden pitäminen muovihuoneessa karaistumisen hidastumisesta huolimatta on järkevää, jotta ne ovat siellä paremmin suojassa syyshalloilta.
Douglaskuusia kasvatettiin kuusen kasvatusohjelman mukaan kastellen niitä yhtä paljon kuin kuusia. Douglaskuuset kasvoivat hitaammin, joten niiden veden tarve oli vähäisempi. Etenkin heinäkuussa ja elokuun alussa kasvualusta oli kosteaa. Douglaskuusella kesällä tehty lievä kuivuuskäsittely on parantanut taimien karaistumista syksyllä (Blake ym. 1979; O’Neill ym. 2001). Liiallinen kasvualustan kosteus douglaskuusen taimillamme on voinut tasaisten lämpötilojen ohella hidastaa douglaskuusen taimien karaistumista verrattuna kuuseen.
Alkuperistä Kouvolan alkuperä oli selvästi kestävin. Douglaskuusen kestävyydet ja LP-käsittelystä saatava hyöty olivat vähäisempiä kuin aiemmissa tutkimuksissa on saatu kuusella. Kuusella LP-käsittely on parantanut silmujen kestävyyttä 3–8 °C ja neulasten ja rangan noin 10 °C alkusyksyn aikana (Luoranen ym. 2008; Riikonen ja Luoranen 2020), kun douglaskuusella silmujen kestävyys parani noin 3 °C ja vain syyskuun alussa ja neulasten ja rangan kestävyys 5–6 °C syys-lokakuussa. Lokakuun lopulla kuusen neulaset ja ranka voivat kestää –35– –50 °C (Riikonen ja Luoranen 2020). Douglaskuusella vain Kouvolan alkuperän LP-käsitellyillä taimilla näiden osien kestävyys oli vähintään –30 °C lokakuun loppupuolella, muiden alkuperien osalta kestävyys oli noin –25 °C. Silmujen kestävyys oli kaikilla alkuperillä alle –20 °C vielä lokakuun lopulla. Syksyllä 2021 Suonenjoen tutkimustarhalla testattiin myös kaksivuotiaiden LP-käsiteltyjen kuusen taimien silmujen kestävyyksiä. Noissa testeissä kuusen silmut kestivät –18 °C jo 27.9. (Riikonen ym. 2023). Tämä viittaa siihen, että douglaskuusen silmut eivät saavuttaisi yhtä hyvää kestävyyttä kuin kotimaisten puulajien taimitarhaolosuhteissa edes LP-käsittelyä käyttäen. Puulajien vertailukokeessa douglaskuusen silmujen kestävyys oli samansuuruinen lokakuussa ja silmunpuhkeamisvaiheessa. Sakain ja Weiserin (1973) tutkimuksessa mereisen douglaskuusen maksimikestävyys oli vain –20 °C. Tulosten perusteella ei voida päätellä sitä, millainen silmujen kestävyys oli keskitalvella. Kokeessamme käytetyt alkuperät ovat niiltä osin kuin lähtöalue on tiedossa Brittiläisestä Kolumbiasta ja on todennäköistä, että niiden maksimikestävyys jää aina heikommaksi kuin kotimaisen kuusen ja männyn. Nicolescun (2021) mukaan douglaskuuset ovat herkimpiä muutaman ensimmäisen vuoden aikana, eikä tulostemme perusteella voida varmuudella sanoa, millainen kestävyys vanhempien taimien silmuilla olisi ja vaarantaako heikompi pakkasenkestävyys taimien jatkokehitystä istutuksen jälkeen.
LP-käsittely oli tehokas keino parantaa douglaskuusen taimien kestävyyttä taimitarhalla varhaisia syyshalloja vastaan kaikilla alkuperillä. LP-käsittelyn käyttö lisää taimien kasvatuskustannuksia, mutta jos samalla voidaan välttää taimien hallakastelusta tulevien kustannuksia sekä syyshalloista aiheutuvia taimitappioita, sen käyttö osana douglaskuusen taimikasvatusta Suomen oloissa on perusteltua.
LP-käsittelyn avulla saavutettiin ruotsalaistutkimuksen (Malmqvist ym. 2017) mukainen douglaskuusen taimien turvallisen pakkasvarastoinnin aloittamiseen vaadittava –20 °C:n raja vain neulasilla. Muiden kasvinosien osalta ei saavutettu edes kanadalaistutkimusten –18 °C:n kestävyysrajaa. Taimien kokonaiskestävyyden kannalta on kuitenkin oleellista, että myös silmut ja ranka kestävät varastoinnin. Silmujen kestävyys jäi kuitenkin kaikilla alkuperillä vaadittua heikommaksi, myös kestävimmäksi todetulla Kouvolan alkuperällä, ja rangat ylittivät vaadittavan kestävyyden vain Kouvolan ja Larch Hillsin alkuperillä. Ilman LP-käsittelyä vain Kouvolan alkuperän neulaset ja rangat olivat riittävän kestäviä. Kokeessa ei määritetty juurten pakkasenkestävyyksiä. Ruotsalaistutkimuksen mukaan alkuperien välillä voi olla eroja myös juurten karaistumisessa (Malmqvist ym. 2017), joten on oletettavaa, että versoiltaan heikommilla alkuperillä juurten kestävyys on ollut meidänkin tutkimuksessamme heikkoa, lisäten noiden alkuperien osalta pakkasvarastoinnin aikaisten juuristovaurioiden riskiä. Tulosten valossa taimien varastoiminen talvella ulkona voi olla riskialtista, ellei taimia suojaavaa lumipeitettä varmisteta.
Silmujen puhkeamisvaiheessa lieviä vaurioita douglaskuusen taimiin tuli jo noin –5 °C:n lämpötiloissa, kun kotoperäiset lajit kestivät useita asteita enemmän pakkasta. Tulos vahvistaa käsitystä, jonka mukaan douglaskuusella on taimivaiheessa suuri riski keväthallan aiheuttamille tuhoille (Nicolescu 2019). LP-käsittelyn on myös todettu aikaistavan sekä kuusen (Konttinen ym. 2003) että douglaskuusen (MacDonald ja Owens 2006) taimien silmun puhkeamista. LP-käsittely muuttaa silmujen rakennetta (Luoranen ja Sutinen 2017) sekä lepotilan purkautumiseen liittyvien geenien aktiivisuutta (Wallin ym. 2017) selittäen todennäköisesti LP-käsiteltyjen taimien aikaisempaa silmujen puhkeamista. LP-käsittely voi siis lisätä taimien hallariskiä keväällä, etenkin, jos taimet ovat olleet talven ulkona. Douglaskuusen alkuperien välillä on myös eroja silmunpuhkeamaisen aikataulussa. Mantereisten alkuperien, kuten kokeissamme olleiden ja Suomessa käytössä olevien, taimien silmut puhkeavat aiemmin kuin mereisten (Malmqvist ym. 2017). Syyksi on arveltu vilutussummien kertymisen eroja, sillä mantereiset alkuperät tarvitsevat vähemmän vilutusta silmujen puhkeamiseen kuin mereiset (Campbell ja Sugano1979). Pakkasvarastossa talven olleilla taimilla riski voidaan välttää ottamalla taimet ulos varastosta vasta hallakauden jälkeen. Toisaalta samalla taimien kasvukausi lyhenee, eikä ole tietoa siitä, kuinka pitkään varastointia voidaan jatkaa ja siten istuttamista kesällä viivästyttää taimien jatkokasvun vaarantumatta. Malmqvistin ym. (2017) tutkimuksessa pakkasvarastoinnin kesto oli neljä kuukautta. Suomessa varastoinnin kestoksi tulee vähintään kuusi, viivästytettäessä istutusta jopa 7–8 kuukautta.
Taimien pakkasvaurioiden määrittäminen oli silmujen ja rangan osalta haastavaa etenkin douglaskuusella lokakuun lopulla. Karaistumisen edetessä rangan ja jossain määrin silmujen vihreä väri vaalenee eikä vaurioitunut osuus muutu välttämättä ruskeaksi, jolloin vaurioituneen ja terveen solukon erottaminen jopa mikroskoopilla on haastavaa. Neulasten osalta vaurioituminen erottui selvemmin. Vauriomäärityksiin voi siis liittyä jonkin verran arviointivirhettä, mutta tulokset antavat suuntaa kestävyydestä ja eri alkuperien eroista, koska hankaluudet määrittämisessä olivat kaikilla alkuperillä ja molemmilla käsittelyillä samanlaisia. Nämä tekijät voivat selittää osin sitä, miksi erot neulasten osalta olivat selkeämmät kuin rangan ja silmujen.
Tutkimuksessa käytettiin pakkasenkestävyyden määrittämiseen visuaalista vaurioiden arviointia, koska halusimme saada selville sekä silmujen, rangan että neulasten kestävyydet. Pakkasenkestävyyden tutkimuksessa käytetään yleisesti myös ionivuoto- ja klorofyllifluoresenssimenetelmiä (Burr ym. 2001). Näissä menetelmissä vauriot määritetään välittömästi altistuksen jälkeen muutamista neulasista koostuvista näytteistä, pienestä osasta rankaa tai versoa (ionivuototesti), mutta visuaalisella menetelmällä voidaan erotella eri kasvinosien mahdollisesti erilainen pakkasenkestävyys. Ionivuototestin käyttökelpoisuus varsinkin hyvin karaistuneilla näytteillä on myös epävarmaa (Sutinen ym. 1992; Luoranen ym. 2004).
Kokeet tehtiin Suonenjoella, joka lienee käytetyille douglaskuusialkuperille varsin pohjoinen paikka kasvukauden pituudeltaan. Jos douglaskuusen viljely yleistyy, on alkuperävalintaan kiinnitettävä huomiota. Tutkimuksessamme oli mukana tietääksemme parhaiten Suomen ilmasto-oloihin sopeutuneet alkuperät, joiden siemeniä oli saatavilla kokeita perustettaessa. Ruotsalainen (2010) toteaa douglaskuusiviljelmien 70 vuoden valtapituuksia tarkastellessaan Larch Hillsin ja Salmon Riverin alkuperien kasvaneen hyvin niin Solbölessä kuin Punkaharjullakin, ja Valemontin alkuperän pärjänneen hyvin Punkaharjulla. Alkuperistä minkään kestävyys taimikasvatuksen aikana ei ollut riittävä riskittömään taimien talvivarastointiin edes LP-käsittelyä käytettäessä. Kouvolan alkuperän alkuperäistä lähtöpaikkaa Pohjois-Amerikassa ei tunneta tarkasti, mutta nyt saatujen tulosten perusteella se on selkeästi Suomen oloihin myös taimivaiheessa paremmin sopeutunut kuin kolme muuta kokeessa mukana ollutta alkuperää. Alkuperän valinnan tärkeyttä korostetaan ruotsalaistutkimuksessa, jossa mereiset ja eteläiset douglaskuusialkuperät saavuttivat pakkasvarastointiin vaadittavan kestävyyden selvästi myöhemmin kuin mantereiset alkuperät (Malmqvist ym. 2017). Koska douglaskuusen kestävyys taimivaiheessa ei ole kovin hyvä edes luontaisilla kasvupaikoillaan (Nicolescu 2019), Suomen oloissa menestyvien alkuperien löytäminen voi olla vaikeaa.
Johtopäätöksenä tutkimuksesta on, että douglaskuusen kestävyys on kotoperäisiä havupuulajeja heikompaa niin syksyllä kuin silmujen puhkeamisvaiheessa. Tulokset osoittavat, että LP-käsittelyllä voidaan parantaa douglaskuusen taimien pakkasenkestävyyttä syksyllä. Saadut kestävyydet eivät kuitenkaan ole riittäviä taimien turvalliseen pakkasvarastointiin. Tämä kasvattaa taimien pakkasvaurioiden riskiä sekä pakkas- että etenkin ulkovarastoinnissa lumettomissa olosuhteissa. Tulostemme valossa taimivaiheen heikko pakkasenkestävyys voi vaikeuttaa kannattavaa douglaskuusen taimituotantoa Suomen oloissa ja olla suuri riski myös taimien maastomenestymiselle ja hallavaurioiden takia taimikoiden laadulle, ainakin keskisen Suomen oloissa.
Selvitys tutkimusdatan, aineistojen ja ohjelmakoodin avoimuudesta
Tutkimusdata on saatavilla sähköpostipyynnöllä jaana.luoranen@luke.fi. Aineistosta piirretyt kuvat, tilastollisen analyysin SAS-koodi sekä estimoidut parametrit on esitetty liitteinä. Tutkimusaineisto oli kertakäyttöinen ja tuhoutui vauriomäärityksissä, eikä ole enää saatavilla. Tutkimus- ja analyysisuunnitelmaa ei ole ennakkorekisteröity.
Kirjoittajien roolit
Kaikki kirjoittajat, Jaana Luoranen (JL), Hanna Ruhanen (HR), Minna Kivimäenpää (MK), Katri Himanen (KH) ja Johanna Riikonen (JR) osallistuivat tutkimuskysymyksen ja tutkimusasetelman muotoiluun, aineiston hankintaan, kirjoituksen kriittiseen muotoiluun hyväksi tieteelliseksi sisällöksi, ovat hyväksyneet julkaistavan version ja ottavat vastuun työn täsmällisyydestä, johdonmukaisuudesta ja laadusta. Lisäksi JL teki aineiston analyysit, tulosten tulkinnan sekä artikkelin tieteellisen kirjoittamisen.
Kiitokset
Kiitämme Sirpa Kolehmaista, Auli Lehtistä, Jenni Toivakkaa, Liisa Kauppista ja Sirpa Mäkistä tutkimuksessa avustamisesta sekä Suonenjoen tutkimustaimitarhan muuta henkilökuntaa taimimateriaalin kasvattamisesta tutkimustamme varten.
Rahoitus
Tutkimus rahoitettiin osana Luonnonvarakeskuksen hankkeita 41007-00100100 (Taimitaito) ja 41007-00231700 (StressTools) sekä Maa- ja metsätalousministeriön rahoittamia MEKANEN- (41007-00188901) sekä Hiilestä kiinni toimenpidekokonaisuuteen kuluvaa PUUVA (41007-00244500) -hankkeita. Viimeksi mainitussa rahoituslähteenä on Euroopan unioni – NextGenerationEU.
Kirjallisuus
Bigras FJ, Ryyppö A, Lindström A, Stattin E (2001) Cold acclimation and deacclimation of shoots and roots of conifer seedlings. Julkaisussa: Bigras FJ, Colombo SJ (toim) Conifer cold hardiness. Kluwer Academic Publishers, Dordrecht, Alankomaat, s. 57–88. https://doi.org/10.1007/978-94-015-9650-3_3.
Blake J, Zaerr J, Hee S (1979) Controlled moisture stress to improve cold hardiness and morphology of Douglas-fir seedlings. For Sci 25: 576–582. https://doi.org/10.1093/forestscience/25.4.576.
Burr K, Hawkins CDB, L’Hirondelle SJ, Binder WD, George MF, Repo T (2001) Methods for measuring cold hardiness of conifers. Julkaisussa: Bigras FJ, Colombo SJ (toim) Conifer cold hardiness. Kluwer Academic Publishers, Dordrecht, Alankomaat, s. 369–402. https://doi.org/10.1007/978-94-015-9650-3_14.
Bärmann L, Kaufmann S, Weimann S, Hauck M (2023) Future forests and biodiversity: effects of Douglas fir introduction into temperate beech forests on plant diversity. For Ecol Manage 545: article id 121286. https://doi.org/10.1016/j.foreco.2023.121286.
Campbell RK, Sugano AI (1979) Genecology of bud-burst phenology in Douglas-fir: response to flushing temperature and chilling. Bot Gaz 140: 223–231. https://doi.org/10.1086/337079.
Di B, Luoranen J, Lehto T, Himanen K, Silvennoinen M, Silvennoinen R, Repo T (2019) Biophysical changes in the roots of Scots pine seedlings during cold acclimation and after frost damage. For Ecol Manage 431: 63–72. https://doi.org/10.1016/j.foreco.2018.04.008.
EPPO Platform of PRAs (2018) Pest categorisation of Ips typographus. EPPO secretariat. https://pra.eppo.int/pra/42a307df-6887-4a20-b5e6-15cd570f9bf4.
Ilmatieteenlaitos, vertailukausi. https://www.ilmatieteenlaitos.fi/ilmastollinen-vertailukausi. Viitattu 15.11.2022.
Jansons Ā, Matisons R, Šēnhofa S, Katrevičs J, Jansons J (2016) High-frequency variation of tree-ring width of some native and alien tree species in Latvia during the period 1965–2009. Dendrochronologia 40: 151–158. https://doi.org/10.1016/j.dendro.2016.10.003.
Konttinen K, Rikala R, Luoranen J (2003) Timing and duration of short-day treatment of Picea abies seedlings. Balt For 9: 2–9.
Konttinen K, Luoranen J, Rikala R (2007) Growth and frost hardening of Picea abies seedlings after various night length treatments. Bal For 13: 140–148.
Lappi J, Luoranen J (2018) Testing the differences of LT50, LD50, or ED50. Can J For Res 48: 729–734. https://doi.org/10.1139/cjfr-2017-0377.
Lehtonen I, Venäläinen A, Gregow H (2020) Ilmastonmuutoksen vaikutukset Suomessa metsänhoidon näkökulmasta. Ilmatieteen laitos. Raportteja 2020:5. https://doi.org/10.35614/isbn.9789523361270.
Lindström A, Stattin E, Gräns D, Wallin E (2014) Storability measures of Norway spruce and Scots pine seedlings and assessment of post-storage vitality by measuring shoot electrolyte leakage. Scand J For Res 29: 717–724. https://doi.org/10.1080/02827581.2014.977340.
van Loo M, Dobrowolska D (2019) History of introducing Douglas fir to Europe. Julkaisussa: Spiecker H, Lindner M, Schuler J (toim) Douglas-fir – an option for Europe. What Science Can Tell Us 9: 21–25.
Luoranen J, Rikala R (2016) Hallakastelun käyttö pakkasvaurioiden estämiseen. Taimiuutiset 2/2016: 26–28. http://urn.fi/URN:NBN:fi-fe2016101125036.
Luoranen J, Sutinen S (2017) Reduced height of short day induced bud scale complex may partly explain early bud burst in Norway spruce seedlings. Silva Fenn 51, article id 7759. https://doi.org/10.14214/sf.7759.
Luoranen J, Repo T, Lappi J (2004) Assessment of frost hardiness of shoots of silver birch (Betula pendula Roth) seedlings with and without controlled freezing exposure. Can J For Res 34: 1108–1118. https://doi.org/10.1139/x03-285.
Luoranen J, Lahti M, Rikala R (2008) Frost hardiness of nutrient-loaded two-year-old Picea abies seedlings in autumn and at the end of freezer storage. New For 35: 207–220. https://doi.org/10.1007/s11056-007-9072-5.
MacDonald JE, Owens JN (2006) Morphology, physiology, survival, and field performance of containerized coastal Douglas fir seedlings given different dormancy-induction regimes. HortScience 41: 1416–1420. https://doi.org/10.21273/HORTSCI.41.6.1416.
MacDonald JE, Owens JN (2010) Physiology and growth of containerized coastal Douglas fir seedlings given different durations of short days to induce dormancy. HortScience 45: 342–346. https://doi.org/10.21273/HORTSCI.45.3.342.
Malmqvist C, Wallertz K, Lindström A (2017) Storability and freezing tolerance of Douglas fir and Norway spruce seedlings grown in mid-Sweden. Scand J For Res 32: 30–38. https://doi.org/10.1080/02827581.2016.1183704.
Malmqvist C, Wallertz K, Johansson U (2018) Survival, early growth and impact of damage by late-spring frost and winter desiccation on Douglas-fir seedlings in southern Sweden. New For 49: 723–736. https://doi.org/10.1007/s11056-018-9635-7.
McCreary DD, Tanaka Y, Lavender DP (1978) Regulation of Douglas-fir seedling growth and hardiness by controlling photoperiod. For Sci 24: 142–152. https://doi.org/10.1093/forestscience/24.2.142.
Mikkonen S, Laine M, Mäkelä HM, Gregow H, Tuomenvirta H, Lahtinen M, Laaksonen A (2015) Trends in the average temperature in Finland, 1847–2013. Stoch Environ Res Risk Assess 29: 1521–1529. https://doi.org/10.1007/s00477-014-0992-2.
Nicolescu V-N (2019) Natural range, site requirements and shade tolerance. Julkaisussa: Spiecker H, Lindner M, Schuler J (toim) Douglas-fir – an option for Europe. What Science Can Tell Us 9: 33–39.
O’Neill GA, Adams WT, Aitken SN (2001) Quantitative genetics of spring and fall cold hardiness in seedlings from two Oregon populations of coastal Douglas-fir. For Ecol Manage 149: 305–318. https://doi.org/10.1016/S0378-1127(00)00564-8.
Partanen J (2004) Dependence of photoperiodic response of growth cessation on the stage of development in Picea abies and Betula pendula seedlings. For Ecol Manage 188: 137–148. https://doi.org/10.1016/j.foreco.2003.07.017.
Repo T, Nilsson J-E, Rikala R, Ryyppö A, Sutinen M-L (2001) Cold hardiness of Scots pine (Pinus sylvestris L.). Julkaisussa: Bigras FJ, Colombo SJ (toim) Conifer cold hardiness. Kluwer Academic Publishers, Dordrecht, Alankomaat, s. 463–493. https://doi.org/10.1007/978-94-015-9650-3_17.
Riikonen J, Luoranen J (2020) An assessment of storability of Norway spruce container seedlings in freezer storage as affected by short-day treatment. Forests 11, article id 692. https://doi.org/10.3390/f11060692.
Riikonen J, Ruhanen H, Luoranen J (2023) Impact of warm spells during late fall and winter on frost hardiness of short-day treated Norway spruce seedlings. For Ecol Manage 542, article id 121105. https://doi.org/10.1016/j.foreco.2023.121105.
Rikala R (2012) Metsäpuiden paakkutaimien kasvatusopas. Metsäntutkimuslaitos. http://urn.fi/URN:ISBN:978-951-40-2359-0.
Ritchie GA, Landis TD, Dumroese RK, Haase DL (2010) The container tree nursery manual – Volume 7. Seedling processing, storage, and outplanting. Chapter 2. Assessing plant quality.
Roques A, Auger-Rozenberg M-A, Capretti P, Sauvard D, La Porta N, Santini A (2019). Pests and diseases in the native and European range of Douglas-fir. Julkaisussa: Spiecker H, Lindner M, Schuler J (toim) Douglas-fir – an option for Europe. European Forest Institute, s. 63–72.
Ruokavirasto 2024. Käyttöaluekartat. Hyväksytyt siemenviljelykset. https://www.ruokavirasto.fi/kasvit/metsapuiden-siemenet-ja-taimet/perusaineisto/kayttoaluekartat/. Viitattu 4.4.2024.
Ruosteenoja K, Markkanen T, Räisänen P (2020) Thermal seasons in northern Europe in projected future climate. Int J Climatol 40: 4444–4462. https://doi.org/10.1002/joc.6466.
Ruotsalainen S (2010) Alkuperän vaikutus puulajin menestymiseen. Sorbifolia 41: 149–172. https://www.dendrologianseura.fi/doc/ruotsalainen_2010.pdf.
Ruotsalainen S, Himanen K, Viherä-Aarnio A, Aarnio L, Haapanen, M, Luoranen J, Matala J, Riikonen J, Uotila K, Ylioja T (2022) Puulajivalikoiman monipuolistaminen metsänviljelyssä: synteesiraportti. Luonnonvara- ja biotalouden tutkimus 24/2022. Luonnonvarakeskus, Helsinki. http://urn.fi/URN:ISBN:978-952-380-394-7.
Sakai A, Weiser CJ (1973) Freezing resistance of trees in North America with reference to tree regions. Ecology 54: 118–126. https://doi.org/10.2307/1934380.
Silander V, Lehtonen J, Nikkanen T (2000) Ulkomaisten havupuulajien menestyminen EteläSuomessa. Metsäntutkimuslaitoksen tiedonantoja 787: 1–127. http://urn.fi/URN:ISBN:951-40-1754-4.
Suomen virallinen tilasto (SVT). Metsänhoito- ja metsänparannustyöt. Luonnonvarakeskus, Helsinki. https://www.luke.fi/fi/tilastot/metsanhoito-ja-metsanparannustyot/metsanhoito-ja-metsanparannustyot-2022. Viitattu 4.4.2024.
Sutinen M-L, Palta JP, Reich PB (1992) Seasonal differences in freezing stress resistance of needles of Pinus nigra and Pinus resinosa: evaluation of the electrolyte leakage method. Tree Physiol 11: 241–254. https://doi.org/10.1093/treephys/11.3.241.
Van Steenis E (1992) Short day treatment of conifer seedlings in British Columbia forest nurseries. Julkaisussa: Landis TD (technical coordinator) Proceedings, Intermountain Forest Nursery Association, 1991 August 12–16, Park City, UT. General Technical Report RM-211: 103–106. Fort Collins, CO, U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. http://www.fcnanet.org/proceedings/1991/vansteenis.pdf.
Venäläinen A, Lehtonen I, Laapas M, Ruosteenoja K, Tikkanen O-P, Viiri H, Ikonen V-P, Peltola H (2020) Climate change induces multiple risks to boreal forests and forestry in Finland: a literature review. Glob Change Biol 26: 4178–4196. https://doi.org/10.1111/gcb.15183.
Viiri H (2020) Parantaako vai heikentääkö metsänhoito metsien kestävyyttä metsätuhoja vastaan? Metsätieteen aikakauskirja, artikkelitunnus 10505. https://doi.org/10.14214/ma.10505.
Wallin E, Gräns D, Jacobs DF, Lindström A, Verhoef N (2017) Short-day photoperiods affect expression of genes related to dormancy and freezing tolerance in Norway spruce seedlings. Ann For Sci 74, article id 59. https://doi.org/10.1007/s13595-017-0655-9.
Weiser CJ (1970) Cold resistance and injury in woody plants. Science 169: 1269–1278. https://doi.org/10.1126/science.169.3952.1269.
47 viitettä.