Fred O. Asiegbu 
Metsäpuiden terveyden suojelu: bioteknologia-ajan haasteita ja mahdollisuuksia
Asiegbu F. O. (2020). Metsäpuiden terveyden suojelu: bioteknologia-ajan haasteita ja mahdollisuuksia. Metsätieteen aikakauskirja vuosikerta 2020 artikkeli 10495. https://doi.org/10.14214/ma.10495
-
Asiegbu,
Helsingin yliopisto, Maatalous-metsätieteellinen tiedekunta, Helsinki
Sähköposti
fred.asiegbu@helsinki.fi
Vastaanotettu 8.12.2020 Hyväksytty 8.12.2020 Julkaistu 22.12.2020
Katselukerrat 7156
Saatavilla https://doi.org/10.14214/ma.10495 | Lataa PDF

 |
(Englanninkielisen alkuperäistekstin käännös: Pekka Nygren, käännöksen tarkistus: Jarkko Hantula)
Metsäpuut ovat tärkeitä paitsi ilmastonmuutoksen vaikutusten lieventämisessä myös suuren ekologisen ja taloudellisen arvonsa takia. Metsäpuut ovat yhtä lailla tärkeitä vaihtoehtoisena bioenergian lähteenä, ja niillä on tärkeä rooli sekä saastumisen vähentämisessä että luonnon monimuotoisuuden ylläpidossa. Metsäpuista saatava puutavara ja siihen liittyvät tuotteet ovat merkittäviä tulolähteitä monille maailman maille. Puutavaran ja metsäntuotteiden kysynnän odotetaan jatkavan kasvuaan ennustettavissa olevassa tulevaisuudessa. Puutuotteiden kestävän tarjonnan tärkeimmät uhat ovat epäsuotuisa ilmasto, hyönteiset ja kasvitaudit. Vaikka metsäpuiden kanssa elävillä mikrobeilla on tärkeä rooli puiden terveyden edistämisessä, siten että ne vahvistavat puiden elinkelpoisuutta ja tuotosta, osa mikrobeista aiheuttaa vakavia kasvitauteja. Viimeisen sadan vuoden aikana on havaittu merkittäviä puiden patogeenisiä infektioita kuten kastanjansurmaa (Cryphonectria parasitica (Murrill) M.E. Barr), hollanninjalavatautia (Ophiostoma Syd. & P.Syd spp.), tammen versopoltetta (Phytophthora ramorum Werres, De Cock & Man in ‘t Veld) ja viime aikoina saarnensurmaa (Hymenoscyphus fraxineus (T. Kowalski) Baral, Queloz & Hosoya). Vuosien kuluessa metsäpatologit ovat edistyneet merkittävästi useiden vaikeiden tautien torjunnassa.
Huolimatta kaikista ponnistuksista puiden tautien leviämisen kontrolloimiseksi, metsätauteja aiheuttavat vieras- ja tulokaslajit muodostavat edelleen uhan puutuotteiden kestävälle tarjonnalle. Nykyisin tarve kasvattaa ja suojella uusiutuvien materiaalien ja bioenergian raaka-aineita sekä ylläpitää puuaineen laatua yhdessä uusien puuperäisten biotuotteiden kasvavan kysynnän kanssa muodostavat merkittäviä haasteita metsien ja puiden terveyden bioteknologiselle tutkimus- ja kehitystyölle. Suomessa yksittäinen tärkeä patogeeni, juurikääpä (Heterobasidion annosum s.l.), aiheuttaa havupuiden vaikean tyvilahotaudin (Kuva 1). Noin 15 % hakkuissa kaadetuista kuusista (Picea abies (L.) H. Karst.) on osin lahonneita ja taloudellisesti vähäarvoisia tämän patogeenin takia (Kuva 2). Juurikääpäinfektiot aiheuttavat Suomessa vuosittain noin 50 miljoonan euron tulonmenetykset ja koko Euroopassa arviolta 800 miljoonan euron tappiot. Sieni esiintyy kaikkialla pohjoisen pallonpuoliskon havumetsissä. Se on ollut vuosia laajan tutkimuksen kohteena.

Kuva 1. Juurikäävän (Heterobasidion annosum s.l.) itiöemä.

Kuva 2. A–B: Juurikäävän (Heterobasidion annosum s.l.) lahottamaa puuainetta ja C: terve puu.
Juurikäävän tiedetään leviävän itiöemistä ilmateitse kulkeutuvien itiöiden laskeutuessa tuoreille kantopinnoille. Kuusella, jolla sieni on Suomessa yleinen, itiöemät kasvavat pääosin juurten alapinnoille, josta ne tulevat esiin ainoastaan puiden kaatuessa juuripaakun kanssa myrskyissä. Tuoreella kantopinnalla rihmastot kasvavat itiöistä alas kantoon ja juuriin ja siirtyvät juuriyhteyksien kautta edelleen lähellä oleviin terveisiin puihin (Kuva 3). Itiöt leviävät pääosin lämpimään kesäaikaan, ja siksi hakkuut on parasta tehdä talvisin.
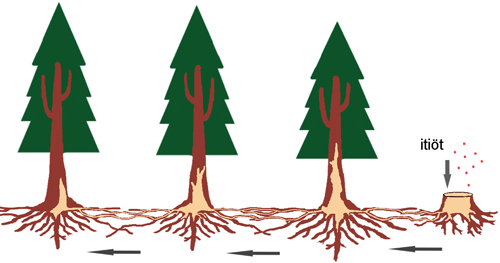
Kuva 3. Kaavio juurikäävän (Heterobasidion annosum s.l.) leviämisestä luonnonmetsässä. Itiöt putoavat äskettäin kaadettujen puiden kannoille, jonka jälkeen ne itävät ja kehittyvät tartuttavaksi rihmastoksi, joka tunkeutuu ja valtaa kannon. Tartunnan saaneesta kannosta ne leviävät edelleen terveisiin lähipuihin juuriyhteyksien kautta.
Päästyään puun sisään juurikääpä pystyy käyttämään hyväkseen kaikkia kolmea puuaineen rakenneosaa: ligniiniä, selluloosaa ja hemiselluloosaa. Sieni hajottaa suuret ligniini- ja selluloosamolekyylit, minkä seurauksena rungon ja juuriston rakenteellinen kestävyys heikkenee ja puunrunko voi murtua melko maltillisissakin tuulissa. Selluloosan hajoaminen vähentää puun arvoa selluloosa- ja paperitehtailla. Hemiselluloosa on tärkeä kemianteollisuuden raaka-aine – esimerkiksi makeutusaineiden ja öljynjalostuksessa käytettävän furfuraalin tuotannossa – joten myös sen hajoaminen vähentää puun teollista arvoa. Näiden juurikäävän kielteisten vaikutusten takia havupuiden raaka-ainearvo alenee sahatavarasta, paperista ja kemikaaleista polttopuuksi. Suomessa käytetään ureaa taudin kemialliseen ja harmaaorvakkasientä (Phlebiopsis gigantea (Fr.) Jülich) biologiseen torjuntaan. Valitettavasti juurikääpäinfektiota vastaan ei ole olemassa 100 % turvallista keinoa. Siksi uusia bioteknologian menetelmiä tarvitaan taudin taltuttamiseksi.
Havupuun ja juurikäävän välisen isäntä–taudinaiheuttajasuhteen kanssa työskentelevä tutkija kohtaa hankalia haasteita. Suuren kokonsa ja pitkän elinkaarensa takia aikuiset havupuut eivät yleensäkään sovellu yksinkertaisin laboratoriomenetelmin tutkittaviksi. Puiden suuren koon takia tutkimusalueen ympäristöä on vaikea kontrolloida. Havupuiden patobiologian tutkimukselle ei myöskään ole olemassa sopivia mallisysteemejä kuten maatalouskasveille (esim. Aradopsis thaliana (L.) Heynh.). Juurikäävästä ei myöskään tunneta yhtään sellaista kantaa, jolla ei olisi taudinaiheuttamiskykyä. Tällä hetkellä DNA:n siirtäminen juurikääpään on erittäin työintensiivistä ja geneettisesti muokattujen kantojen eloonjäänti on vähäistä ja satunnaista, mikä tekee geenien toiminnalliseen poistamiseen perustuvat tutkimukset hankaliksi. Yhdelläkään Pinaceae-heimon havupuulajilla ei tunneta juurikäävälle täysin vastustuskykyistä genotyyppiä. Puut aiheuttavat lisähaasteita kokonsa, pitkän elinkaarensa, pitkien tautijaksojen, suuren genomin ja mutanttilinjojen puutteen takia.
Juurikäävän ja muiden puutautien tutkimuksen haasteellisuudesta huolimatta on olemassa joitakin menestystarinoita ja uusia edistysaskeleita. Hyvä esimerkki on kastanjansurman torjunta hypovirulentin Cryphonectria parasitica -isolaatin avulla, jolla on symbionttinen sienivirus. Myös muuntogeenisen Amerikan kastanjan (Castanea dentata (Marshall) Borkh.), jolla on oksalaatin hapetusgeeni, on osoitettu olevan vastustuskykyinen kastanjansurmalle. Biologista torjuntaa on käytetty myös muiden metsäpuiden tautien hallintaan. Hollanninjalavatautia on kontrolloitu injektoimalla Verticillium albo-atrum Reinke & Berthold -lakastesienen kantaa WCS850 (kauppanimenä DutchTrig) puiden putkiloihin. Myös havupuiden tyvilahoa on torjuttu biologisesti Rotstop-harmaaorvakkavalmisteen avulla.
Kuusen juurikäävänkestävyyden on oletettu olevan monien geenien (kvantitatiivisesti eli polygeenisesti) ohjauksessa, koska se on nekrotrofinen taudinaiheuttaja. Kvantitatiivinen vastustuskyky tarkoittaa epätäydellistä fenotyyppistä vastustuskykyä, joka voi johtua useiden samaan ominaisuuteen vaikuttavien geenien vaihtelusta. Kvantitatiivisen vastustuskyvyn tiedetään olevan kestävää. Vastaavasti kvalitatiivinen eli täydellinen vastustuskyky on yhden geenin ohjaama ja sitä havaitaan yleensä biotrofisia patogeenejä vastaan.
Aiemmassa tutkimuksessamme kuusi- ja mänty-yksilöiden (Pinus sylvestris L.) taudin- ja lahoamisen vastustuskyvyssä havaittiin laajoja geneettisiä eroja, joita selitti vaihtelu esimerkiksi fenoleiden ja terpeenien kokonaismäärässä sekä useiden flavonoidien biosynteesituotteita (esim. kateenit) koodaavien fenyylipropanoidireitin geenien lisääntynyt transkriptio. Alustavissa tutkimuksissamme on tunnistettu myös uusia männyn antimikrobiaalisia peptidejä (Sp-AMP), joilla on vahva juurikäävän itiöiden itämistä ja kasvamista estävä vaikutus. Tämä on tärkeä edistysaskel taistelussa tätä patogeeniä vastaan. Löydetty Sp-AMP-peptidi on nimetty PR-19:ksi ja se edustaa uutta antimikrobisten proteiinien ryhmää. Toinen mielenkiintoinen havainto laboratoriossamme on, että muuntogeeninen tupakka (Tabacum nicotianum Berchtold & Opiz.), jolla on toimiva Sp-AMP, on vastustuskykyinen Botrytis cinerea -harmaahomesienelle. Toiset tutkimusryhmät ovat raportoineet tutkimuksia mykovirusten käytöstä juurikäävän torjunnassa.
Kamppailussa metsäpuiden tauteja vastaan odotetaan uusia edistysaskeleista, jotka perustuvat bioteknologisen tiedon ja työkalujen sekä läheisten tieteenalojen, kuten puiden genetiikan, genomiikan, genomisen valinnan ja resistenssijalostuksen hyödyntämiseen. Globaali ympäristön- ja ilmastonmuutos sekä tulokas- ja vierasperäiset tuhohyönteiset ja patogeenit aiheuttavat painetta lisätä jalostustyöhön uusia tavoitteita, kuten tautien vastustuskyvyn. Siten puiden kestävyyttä ilmastonmuutosta ja kuivuutta vastaan ja vastustuskykyä hyönteisille ja patogeeneille ei voi jättää huomiotta.
Perinteinen puiden jalostustyö vaatii aikaa ja resursseja, koska jalostussykli voi olla vuosikymmenten pituinen. Jalostustyön nopeuttamiseksi on puiden fenotyyppien arviointi ja valinta tehtävä mahdollisimman varhaisessa vaiheessa. Siksi tarvitaan tutkimusta puiden eloonjäämiseen ja kasvuun sekä uusien tautien vastustuskykyyn vaikuttavien ominaisuuksien välisten korrelaatioiden tunnistamiseksi, jotta ne voidaan ottaa mukaan jalostustyöhön. Tämä parantaisi taimien genomisen valinnan käyttömahdollisuuksia, ja sitä kautta jalostussykliä voitaisiin nopeuttaa ja vähentää jälkeläiskokeita. Uudet tekniset edistysaskeleet ovat merkittävästi nopeuttaneet metsäpuiden jalostettujen genomiresurssien saatavuutta, mikä edesauttaa genomisten ennusteiden tekoa suurigenomisista havupuista kuten kuusista ja männyistä. Näillä uusilla metsäpuiden bioteknologian ja jalostuksen edistysaskeleilla voi olla merkittävä rooli vastattaessa metsäteollisuuden tulevaisuuden haasteisiin varmistamalla pitkän ajan kestävyys ja vastustuskyky bioottisille ja abioottisille stressitekijöille.
Bioteknologian viimeaikaiset edistysaskeleet tarjoavat mahdollisuuksia käyttää hyväksi genomisia työkaluja juurikäävän ja muihin metsäpuiden taudinaiheuttajien tutkimukseen. Tämä edistää merkittävästi kykyä torjua leviäviä ja uusia tuhohyönteisiä ja tauteja. Genomiikka edesauttaa tunnistamaan elintärkeitä puuaineen ominaisuuksia, jotka vaikuttavat esimerkiksi tautien vastustuskykyyn (Kuva 4). Niitä ei yleensä oteta huomioon perinteisessä jalostustyössä. Koko genomin assosiaatiotutkimus (Genome wide association study, GWAS) helpottaa yhden nukleotidin polymorfismien (single nucleotide ploymorphism, SNP) etsintää genomeista. Ne auttavat löytämään tautiherkkyyteen ja vastustuskykyyn liittyvää vaihtelua. Geneettisiä ominaisuuksia, jotka ovat yleisempiä vastustuskykyisissä kuin tautiherkissä puiden genotyypeissä, pidetään assosioituneina. Genomiset valintatyökalut ja niihin liittyvät tilastomallit helpottavat edelleen geenitunnisteiden ja fenotyyppien tehokasta havaitsemista. Genomisen valinnan tai ennustamisen hyväksikäytön voi odottaa johtavan lyhentyvään jalostussykliin sekä lisääntyvään ja tehokkaaseen valintatulosten löytymiseen.
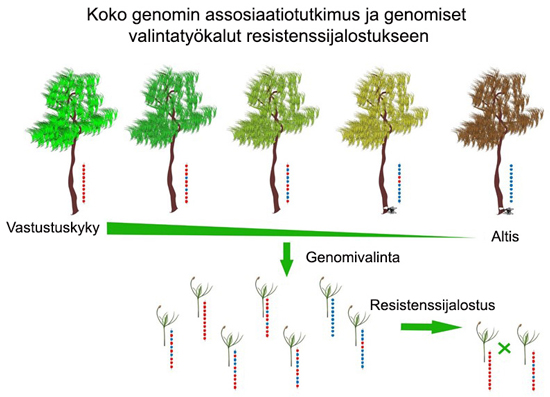
Johtopäätöksenä voidaan todeta, että kemialliset ja biologiset torjuntakeinot ovat lyhytaikaisia tapoja kamppailla puiden infektioita ja tauteja vastaan. Pitkäaikainen, kestävä vastustuskyky saavutetaan vain genetiikan avulla.
Kirjallisuutta
Asiegbu F.O., Adomas A., Stenlid J. (2005). Conifer root and butt rot caused by Heterobasidion annosum (Fr.) Bref. s.l.. Molecular Plant Pathology 6(4): 395–409. https://doi.org/10.1111/j.1364-3703.2005.00295.x.
Canadell J.G., Raupach M.R. (2008). Managing forests for climate change mitigation. Science 320(5882): 1456–1457. https://doi.org/10.1126/science.1155458.
Gauthier S., Bernier P., Kuuluvainen T., Shvidenko A.Z., Schepaschenko D.G. (2015). Boreal forest health and global change. Science 349(6250): 819–822. https://doi.org/10.1126/science.aaa9092.
Haapanen M., Hynynen J., Kilpeläinen M.L. (2016). Realised and projected gains in growth, quality and simulated yield of genetically improved Scots pine in southern Finland. European Journal of Forest Research 135: 997–1009. https://doi.org/10.1007/s10342-016-0989-0.
Isik F. (2014). Genomic selection in forest tree breeding: the concept and an outlook to the future. New Forests 45: 379–401. https://doi.org/10.1007/s11056-014-9422-z.
Kashif M., Jurvansuu J., Vainio E.J., Hantula J. (2019). Alphapartitiviruses of Heterobasidion wood decay fungi affect each other’s transmission and host growth. Frontiers in Cellular and Infection Microbiology 9 article 64. https://doi.org/10.3389/fcimb.2019.00064.
Kovalchuk A., Keriö S., Jaber E., Oghenekaro A., Raffaello T., Asiegbu F.O. (2013). Antimicrobial defences and resistance of forest trees: challenges and perspectives in a genomic era. Annual Review of Phytopathology 51: 221–244. https://doi.org/10.1146/annurev-phyto-082712-102307.
Kovalchuk A., Zeng Z., Ghimire R.P., Kivimäenpää M., Raffaello T., Liu M., Mukrimin M., Kasanen R., Sun H., Julkunen-Tiitto R., Holopainen J.K., Asiegbu F.O. (2019). Dual RNAseq analysis provides new insights into interactions between Norway spruce and necrotrophic pathogen Heterobasidion annosum s.l. BMC Plant Biology 19 article 2. https://doi.org/10.1186/s12870-018-1602-0.
Mukrimin M., Kovalchuk A., Neves L.G., Jaber E., Haapanen M., Kirst M., Asiegbu F.O. (2018). Genome-wide exon-capture approach identifies genetic variants of Norway spruce genes associated with susceptibility to Heterobasidion parviporum infection. Frontiers in Plant Science 9 article 793. https://doi.org/10.3389/fpls.2018.00793.
Neale D.B., Wegrzyn J.L., Stevens K.A., Zimin A.V., Puiu D., Crepeau M.W., Cardeno C., Koriabine M., Holtz-Morris A.E., Liechty J.D., Martínez-García P.J., Vasquez-Gross H.A., Lin B.Y., Zieve J.J., Dougherty W.M., Fuentes-Soriano S., Wu L.-S., Gilbert D., Marçais G., Roberts M., Holt C., Yandell M., Davis J.M., Smith K.E., Dean J.F.D., Lorenz W.W., Whetten R.W., Sederoff R., Wheeler N., McGuire P.E., Main D., Loopstra C.A., Mockaitis K., deJong P.J., Yorke J.A., Salzberg S.L., Langley C.H. (2014). Decoding the massive genome of loblolly pine using haploid DNA and novel assembly strategies. Genome Biology 15 article R59. https://doi.org/10.1186/gb-2014-15-3-r59.
Nystedt B., Street N.R., Wetterbom A., Zuccolo A., Lin Y.C., Scofield D.G., Vezzi F., Delhomme N., Giacomello S., Alexeyenko A., Vicedomini R., Sahlin K., Sherwood E., Elfstrand M., Gramzow L., Holmberg K., Hällman J., Keech O., Klasson L., Koriabine M., Kucukoglu M., Käller M., Luthman J., Lysholm F., Niittylä T., Olson A., Rilakovic N., Ritland C., Rosselló J.A., Sena J., Svensson T., Talavera-López C., Theißen G., Tuominen H., Vanneste K., Wu Z.Q., Zhang B., Zerbe P., Arvestad L., Bhalerao R., Bohlmann J., Bousquet J., Garcia Gil R., Hvidsten T.R., de Jong P., MacKay J., Morgante M., Ritland K., Sundberg B., Thompson S.L., Van de Peer Y., Andersson B., Nilsson O., Ingvarsson P.K., Lundeberg J., Jansson S. (2013). The Norway spruce genome sequence and conifer genome evolution. Nature 497(7451): 579–584. https://doi.org/10.1038/nature12211.
Sooriyaarachchi S., Jaber E., Suarez Covarrubias A., Ubhayasekera W., Asiegbu F.O., Mowbray S. (2011). Expression and β-glucan binding properties of Scots pine (Pinus sylvestris L.) antimicrobial protein (Sp-AMP). Plant Molecular Biology 77(1–2): 33–45. https://doi.org/10.1007/s11103-011-9791-z.
Lähetä sähköpostiin